The hSK4 (KCNN4) isoform is the Ca2؉-activated K؉ channel (Gardos channel) in human red blood cells Joseph F. Hoffman*†, William Joiner*‡, Keith Nehrke§, Olga Potapova*¶, Kristen Foye*ʈ, and Amittha Wickrema**
*Departments of Cellular and Molecular Physiology and Pharmacology, Yale University, New Haven, CT 06520; §Department of Medicine, University ofRochester Medical Center, Rochester, NY 14642; and **Section of Hematology͞Oncology, University of Chicago, Chicago, IL 60637
Contributed by Joseph F. Hoffman, April 21, 2003
The question is, does the isoform hSK4, also designated KCNN4,
cells allow us to study purely erythroid developmental stages free
represent the small conductance, Ca2؉-activated K؉ channel (Gar-
of contamination from white cells or platelets. This conclusion
dos channel) in human red blood cells? We have analyzed human
is based on ‘‘ profiling,’’ by which we mean that the progenitor
reticulocyte RNA by RT-PCR, and, of the four isoforms of SK
cells were found to contain the message for the 2 and not the
channels known, only SK4 was found. Northern blot analysis of
1 isoforms of the  subunit of the Na pump in contrast to a
purified and synchronously growing human erythroid progenitor
white cell and platelet library that contained the message for the
cells, differentiating from erythroblasts to reticulocytes, again
1 but not the 2 isoforms (20). Our studies using progenitor
showed only the presence of SK4. Western blot analysis, with an
cells indicate that SK4 is the isoform that subserves the functions
anti-SK4 antibody, showed that human erythroid progenitor cells
of the Gardos channel in human red blood cells. This identifi-
and, importantly, mature human red blood cell ghost membranes,
cation is based on RT-PCR, Northern and Western blotting of
both expressed the SK4 protein. The Gardos channel is known to
progenitor cells, and Western analysis of human red blood cell
turn on, given inside Ca2؉, in the presence but not the absence of
ghosts. We also found that SK4, when heterologously expressed
external K؉
in Chinese hamster ovary (CHO) cells, displays functional
o and remains refractory to Ko added after exposure to inside Ca2؉. Heterologously expressed SK4, but not SK3, also
characteristics of the Gardos channel similar to those seen in
shows this behavior. In inside–out patches of red cell membranes, the open probability (Po) of the Gardos channel is markedly reduced when the temperature is raised from 27 to 37°C. Net K؉ efflux of intact red cells is also reduced by increasing temperature, Reticulocyte and Erythroid Progenitor Cell RNA. This study used
samples of RNA taken from the same preparations of reticulo-
as are the Po values of inside–out patches of Chinese hamster ovary
cytes and human erythroid progenitor cells purified and cultured
cells expressing SK4 (but not SK3). Thus the envelope of evidence
as described previously (20). All of these preparations were
indicates that SK4 is the gene that codes for the Gardos channel in
found to be free of leukocyte and platelet contamination based
human red blood cells. This channel is important pathophysiologi-
on the criterion of ‘‘ profiling,’’ as previously established (20). cally, because it represents the major pathway for cell shrinkage
RT-PCR was carried out by using the SK isoform specific
via KCl and water loss that occurs in sickle cell disease.
primers given in Table 1. Note that one primer pair, labeled
SK1–3, was designed to detect these three channel isoforms but
Four isoforms (SK1–4) of the small conductance Ca2ϩ- not SK4. To confirm that primers were capable of amplifying a
activated Kϩ channel have been identified (1–5). These
specific product, RT-PCR was performed with each set of
channels, with acronyms small conductance (SK), intermediate
primers and plasmids individually encoding each of the channel
conductance (IK), and potassium channel calcium-activated
isoforms. In each case, a product of the expected size was
intermediate͞small conductance subfamily N (KCNN), are
amplified with Ͼ96% sequence identity.
highly conserved Ca2ϩ-activated inward rectifiers (see refs. 1–5).
Although KCNN is the notation assigned by GenBank (6), we
Northern Blotting. Northern blotting was carried out as described
use herein the SK notation and report studies that deal mainly
before (20). DNA probes were derived from the following
with the human isoform, i.e., hSK4, recognizing that there is a
gel-purified DNA templates: a 436-bp BamHI–HindIII fragment
parallel with the rat, rSK4, and mouse, mSK4, isoforms (7). We
of rat SK1 (rSK1); a 582-bp EcoRV–NarI fragment of rat SK2
are principally concerned with SK4 because of its putative
(rSK2); a 710-bp SmaI–SmaI fragment of rat SK3 (rSK3); and
identification as the Ca2ϩ-activated Kϩ channel, referred to as
a 723-bp SphI–SfiI fragment of human SK4 (hSK4). These DNA
the Gardos channel (8), in human red blood cells. Activation of
fragments were radiolabeled by using a Prime-It II kit (Strat-
the channel results in a marked hyperpolarization of the mem-
agene) and 32P-dCTP and purified on G-50 Spin Columns
brane accompanied by shrinkage of the cell due to the loss of KCl
(Roche Applied Science, Indianapolis) to achieve a specific
and water. The main reason the Gardos channel has been
activity of Ϸ109 dpm͞g. As a positive control, probes were
assigned to the SK4 gene in human red blood cells is because of
also hybridized with a human multiple tissue Northern blot
parallels in its electrophysiological characteristics (7, 9, 10)
between the intact cell and the expressed channel as well as its
pharmacological sensitivities; the channel is inhibited by charyb-
Western Blotting. The anti-mouse SK4 (mSK4͞IKCa) antibody (21)
dotoxin (11) and clotrimazole (CLT) (12) but not by apamin
was custom produced by Research Genetics, a division of Invitro-
(13). The SK4 expressed in different cell types shares this
inhibitory profile (1, 7). In sharp contrast, expressed SK1, 2, or
3 channels are inhibited by apamin (14–17) but not by CLT (18).
Abbreviations: CLT, clotrimazole; CHO, Chinese hamster ovary.
Obviously, it is necessary to go beyond the foregoing correlations
†To whom correspondence should be addressed at: Department of Cellular and Molecular
of channel characteristics to establish which isoform(s) of the SK
Physiology, Yale University School of Medicine, 333 Cedar Street, New Haven, CT 06520-8026. E-mail: [email protected].
family are actually found in human red blood cells.
‡Present address: Department of Neuroscience, Howard Hughes Medical Institute, 232
Our approach exploits the use of human erythroid progenitor
Stemmler Hall, University of Pennsylvania, Philadelphia, PA 19104.
cells in which we have previously determined the subunit types
¶Present address: Department of Molecular Biophysics and Biochemistry, Yale University,
and isoform composition of the endogenously expressed Naϩ
pumps (19, 20). Our particular preparations of the progenitor
ʈPresent address: Genaissance Pharmaceuticals, New Haven, CT 06511. 7366 –7371 ͉ PNAS ͉ June 10, 2003 ͉ vol. 100 ͉ no. 12
www.pnas.org͞cgi͞doi͞10.1073͞pnas.1232342100
Table 1. PCR primer pair sequences (all designed against human
where they were Ca2ϩ-dependent and displayed the expected
sequences) that were selected to have distinct and high
unitary conductance characteristics (1). specificity for each of the indicated SK isoforms of the
The pipette solution for whole-cell experiments consisted of
Ca؉؉-activated K؉ channel
(in mM): 30 KCl, 100 K-gluconate, 5 EGTA, and 10 Hepes (pH
7.2). This solution was supplemented with 4.27 or 4.74 mM CaCl2
to achieve free Ca2ϩ concentrations of 1.0 or 3.0 M, respec-
tively. All whole-cell recordings were performed by using 200-ms
ramps from Ϫ120 to ϩ80 mV from a holding potential of Ϫ70
mV. Current densities were measured for each cell by dividing
the current amplitude at 60 mV by the capacitance.
For inside-out patch recordings of stably transfected cells,
pipettes were filled with (in mM): 30 KCl, 100 K-gluconate, 1
MgCl2, and 10 Hepes (pH 7.2), and the cytoplasmic side of the
membrane was perfused with (in mM): 30 KCl, 100 K-gluconate,
5 EGTA, 10 Hepes (pH 7.2), and 4.27 CaCl2 (1 M free Ca2ϩ).
In some experiments, lyophilized thioredoxin peroxidase (23),
i.e., calpromotin (24, 25), was dissolved in the latter solution at
a concentration of 10 M and used to perfuse excised inside-out
patches. All patch recordings were made over a period of 30–60
s at a holding potential of Ϫ80 mV, first at 25°C and subse-
quently, after rapidly switching (Ͻ10 s), at a controlled bath
temperature of 35°C in a temperature-controlled 35-mm tissue
culture cup. Electrical heating was mediated via a Pt͞Ir oxide
film on the outside of the cup (Bioptechs, Butler, PA). and 86Rb؉ Fluxes. Blood was drawn into heparin from
In the case of the SK1–3 (i.e., SK1, SK2, and SK3) primer set, M ϭ A or C; R ϭ
normal volunteers and used without delay. Net Kϩ effluxes were
measured by incubating red cells in a low Kϩo medium for various
time periods. The Gardos channel (i.e., Ca2ϩ-activated Kϩ
gen. The antibody was directed toward an mSK4-specific peptide
channel) was activated, in the presence of Ca2ϩ
with the amino acid sequence RQVRLKHRKLTEQVNSMVD.
the divalent ionophore, A23187, or energy depletion or both, as
Pellets of 7- and 13-day-old cultures of human erythroid progenitor
indicated below. The experimental protocols used were varia-
cells (see ref. 20) as well as human brain, kidney, and parotid tissues
tions on those described by others (26, 27). Because details of the
were prepared by previously described methods (see ref. 21).
protocols varied, we have adopted abbreviations for some con-
Human red blood cell ghosts were prepared from heparinized
stituents that were common to many of the solutions used. Thus,
peripheral blood by hypotonic lysis, as described (22), and were
H is Hepes buffer, C is CaCl2, M is 0.2 mM MgCl2, A is 10 M
frozen, thawed, and washed before use.
A23187, V is 1.0 mM orthovanadate, I is 10 mM inosine, IA is
Approximately10 g of crude protein was separated by two-
6.0 mM iodoacetamide, and CLT is 10 M chlotrimazole. It
phase Tricine polyacrylamide gel electrophoresis (10% T͞6% C
should be understood that V is used to inhibit the Ca2ϩ pump,
resolving layer, 4% T͞3% C stacking layer) and transferred onto
thereby allowing for the accumulation of (Cai ) necessary to
poly(vinylidene difluoride) membrane (BioDyne PVDF, Pall
activate the Gardos channel. The combination of I plus IA is
Filtration, General Electric) in buffer containing 10 mM 3-
used to deplete the cells of energy (ATP) that could interfere
[cyclohexylamino]-1-propanesulfonic acid (CAPS) adjusted to
with the activation of the Gardos channel. In all experiments,
pH 11 and 10% methanol. Blotting was then carried out as
final incubations were carried out in the absence and presence
described (21). To assess specificity, the mSK4 antibody was
of CLT, and all samples taken from suspensions where CLT was
preincubated for 1 h with a 50-fold molar excess of competitor
absent were mixed before centrifuging with a stop solution
containing CLT. This procedure provided the time resolution for
peptide corresponding to the epitope recognition sequence
rapid fluxes. Hematocrits for net Kϩ measurements varied from
11% to 20%. The pH of all solutions was Ϸ7.4 at 37°C. Changing
the temperature from 37 to 27°C lowers the pH by Ϸ0.1 pH unit,
Stable SK-Expressing Cell Lines. CHO cells stably expressing hSK4
without appreciable effect on the Kϩ flux (28). For Table 2, the
channels as described (1) were used in this study. Another CHO
following experimental conditions were used. Experiment A:
cell line stably expressing rSK3 was established by similar means,
cells were washed with a solution containing (in mM) 40 NaCl,
except that the cDNA was carried in the plasmid pcDNA 3.1 Zeo
90 NaSCN, 2 KCl, M, and 20 H. The cells were then energy
(from Guy Moss, University College, London), and Zeocin
depleted by incubation in this solution together with 1.3 mM C,
(Invitrogen) was used to select stable transfectants. Stably
I, IA, and V for 25 min at 37°C. The suspension was then split,
transfected survival colonies were sorted by FACS as described
with half incubated at 27°C and the other half at 37°C. After 3
(1). A single clonal population of cells was used for each cell line.
min, samples were taken at 0, 15, 30, 45, and 60 min. Experiment
B: cells were washed in the presence or absence of SCNϪ in
Electrophysiology of SK Channels. Single-channel recordings were
solutions containing (in mM): either 40 NaCl ϩ 90 NaSCN or
performed by using the methods described (1). For whole-cell
130 NaCl together with 0.5 KCl, 30 H, and M. After washing, the
recordings, the standard bath solution (5 mM Ko) consisted of
cells were suspended, respectively, in each of these solutions
(in mM): 140 NaCl, 1.0 CaCl2, 5 KCl, 29 glucose, 25 Hepes (pH
together with the additions of 1.3 mM C, I, IA, and V. The cells
7.4). Bath composition for ion substitution experiments was
were then energy depleted by incubation in these solutions for 25
min at 37°C. Then each solution was split, with half incubated at
0 mM Kϩo. It should be understood that the Kϩ channels studied
27°C and the other half at 37°C. After 3 min, aliquots were taken
in CHO cells were seen not in control cells but only in cells that
over a 30-min period for determination of Kϩ
PHYSIOLOGY
had been transfected with either the rSK3 or hSK4 isoforms,
cells were energy depleted by incubation for 3 h in a solution
PNAS ͉ June 10, 2003 ͉ vol. 100 ͉ no. 12 ͉ 7367 Table 2. The effect of temperature on the net efflux of K؉ from human red blood cells after activation of the Gardos channel by various means
The SK4 isoform of the Ca2ϩ-activated Kϩ channel is present in human
The effluxes of Kϩ were measured at either 27 or 37°C (see Methods for
reticulocytes. PCR products were obtained with the isoform-specific primers
details) and estimated from the rate of increase in Kϩ
defined in Table 1. Single-stranded cDNA derived from reticulocytes (see
o medium. The cells were treated with V (except Experiment C) to
Methods) was used as template. Lanes 1 and 2 had, respectively, 25- and 4-l
inhibit Caϩϩ efflux via the Caϩϩ pump. In addition, the cells were energy
samples applied to the gel. Lane 3 is a water control. The mass ladder (in base
depleted in Experiments B and C but not in A, in which they were normal. Kϩ
pairs) is shown at left (Life Technologies, Grand Island, NY). The expected
efflux from cells were begun in Experiments A and B after a 3- to 5-min
product size was 1,767, and a product of approximately this size was found in
equilibration at their respective temperatures, and in Experiment C, after
lanes 1 and 2. The product was sequenced in this and other analyses and was
addition of the ionophore A23187 (A23). The Kϩ
shown to have Ͼ98% identify with the expected sequence for SK4.
o (mM) values refer to time zero. The effluxes of Ki were also estimated
from the rate of appearance of medium 86Rb from cells preloaded with 86Rb(Experiment D). The cells were treated with V to inhibit Caϩϩ efflux via the
analogous results with the second set of SK4 primers given in
Caϩϩ pump. In addition, the cells were energy depleted. 86Rb effluxes, from
Table 1 (data not shown). Use of any of the other isoform
cells having been washed in the cold, were begun after a 3- to 5-min equili-bration of the cell suspensions at their respective temperatures. All effluxes
primers listed in Table 1 for SK1, 2, or 3 was negative with regard
were carried out in the absence and presence of CLT and presented as the
to their specific presence in these RNA preparations. When
CLT-sensitive flux (⌬CLT). The CLT-insensitive efflux was in all cases Ͻ5–10% of
products did appear, they were neither of the expected size nor
efflux (Experiments A–C) appeared to be
did they display any sequence homology with the isoform being
exponential in all cases, the outward rate constant, °k⌬CLT
tested. Because we used preparations of reticulocytes that were
mated from the initial rate, as explained in Methods. The values of °k⌬CLT
free from white cell and platelet contamination by the criterion
Experiment D represent the means Ϯ SEM, where n ϭ 4.
of ‘‘ profiling’’ (20), the results strongly indicated that SK4 was
the isoform responsible for the Gardos channel in human
containing (in mM): 10 NaCl, 50 KCl, 60 KSCN, 1.0 Na
erythrocytes. However, there is a caveat to this interpretation,
30 H, I, IA, and V. The cells were washed and resuspended in
because SK4 has also been identified as a constituent of human
the following solution (in mM): 60 NaCl, 60 NaSCN, 0.4 KCl, 30
lymphocytes (3). Thus, although contamination of our prepara-
H, M, I, IA, and V together with 50 M C. After 3 min at either
tions is improbable, it must remain a possibility.
27 or 37°C, the flux was initiated by addition of A with samples
We next turned to our preparations of human erythroid
taken at 0, 45, 90, 135, and 180 s. Experiment D: Cells loaded
progenitor cells, because, as shown before (20), an erythroid ‘‘
with 86Rb by incubation for 3 hr in (in mM): 30 KCl, 90 KSCN,
profiling’’ was also a characteristic of these differentiating cells.
20 H and M. Cells were then resuspended in this solution
Fig. 2 shows the results of Northern analysis of these progenitor
together with 1.3 mM C, I, IA, and V and incubated at 37°C for
cells from days 7–12 of culture. The evidence presented in Fig.
15 min before washing in the cold. Cells were resuspended at
2 shows that SK4 is present, and there is a clear tendency for its
27°C and 37°C and after 3 min samples were taken at 0, 5, 10, 15,
expression level to increase with increasing maturation. The
positive control blots are given on the right side of Fig. 2 and
Calculation of 86Rb rate constants were the same as previously
made use of mRNA from human placental tissue, which is known
described (29). Hematocrits (Hcts) were calculated by using the
(17) to contain the SK4 isoform (1, 17). Although we saw no
equation, Hct ϭ (SWB Ϫ So)͞(Si Ϫ So), where SWB is the whole
evidence by Northern analysis for SK1 or SK3, some blots
blood, cellular (Si), or supernatant (So) concentration of Kϩ. The
indicated that hSK2 might be present. This was not pursued
outward rate constants (hrϪ1) calculated for the results in Table
because of failure to find the SK2 isoform by RT-PCR and
because the Gardos flux in human erythrocytes is not inhibited
Ro͞[Kϩ]i, where Ro (in mM͞unit time) is the initial rate assuming
by apamin (13). Thus we conclude from Northern analysis that
the curves are single exponentials, and [Kϩ]i is in mmol͞l cell
water. The percent water is taken from the difference in wet and
dried weights of packed cells. CLT, inosine, and iodoacetamide
were obtained from Sigma; A23187 from Calbiochem; and
1,2-bis(2-aminophenoxy)ethane-N,N,NЈ,NЈ-tetraacetate or
-tetraacetic acid–acetoxymethyl ester (BAPTA-AM) from Mo-
lecular Probes. All other chemicals, wherever possible, were of
Results Our prime aim in undertaking the studies reported here was to
Northern blots probed for the mRNA encoding the Ca2ϩ-activated Kϩ
identify the gene that codes for the Gardos channel (or Ca2ϩ-
channel isoform, SK4, using RNA prepared from cultured human erythroid
dependent K permeability) of human red blood cells. We first
progenitor cells at different stages of maturation (see Methods for details).
screened, by RT-PCR, our previous preparations of RNA ex-
Also shown is the positive control blot for SK4 (Right), where H is heart; B,
tracted from human reticulocytes (19) by use of the primer sets
brain; P, placenta; L, lung; Li, liver; S, skeletal muscle; K, kidney; and Pa,pancreas, all from human mRNA. These results parallel our previous finding
listed in Table 1, to establish which isoforms of the SK (or
that SK4 is present in reticulocytes. Note that SK4 is present in the progenitor
KCNN) channel family were present. As is evident in Fig. 1, the
cells at day 7 increasing with differentiation through day 12. The transcript
isoform SK4 is present in these preparations. We also obtained
7368 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.1232342100
Western blots showing that the protein for the Gardos channel
isoform, SK4, is present in cultured human erythroid progenitor cells and inghost membranes made from mature human red blood cells. The antibodywas prepared against an SK4-specific peptide and used as described in Meth-ods. The two positive controls are human parotid gland (P) and kidney (K) withthe negative control being brain (B). It is clear that a band of the appropriatemolecular weight is present in the human erythroid progenitor cells as theymature from days 7 to 13. It is also evident that the SK4 band is present inhuman red cell ghost membranes (RBC). The decrease in the blot intensity ofthe D13 band compared with D7 is primarily due to the decreased proteincontent (cell number) of cells loaded onto the gel. The slight variation in themolecular weights of SK4 bands seen in the progenitor cells, relative to theother bands, may be due to posttranslational modification or higher saltconcentration in the loading mixture. It should also be mentioned that, exceptin the parotid lane, there are higher molecular weight bands (not shown) thatin each case react with the antibody. Importantly, preincubation of theantibody with purified peptide that contains the antigenic epitope producesa complete loss of reactivity in all lanes except in brain, where it is muchreduced, and in ghosts, where it is only faintly present in the highest molecularweight bands (data not shown).
differentiating human erythroid progenitor cells expresses only
We also performed a Western blot analysis, the results of
which are shown in Fig. 3. By using an antibody directed against
a peptide whose sequence is derived from mSK4, we find that a
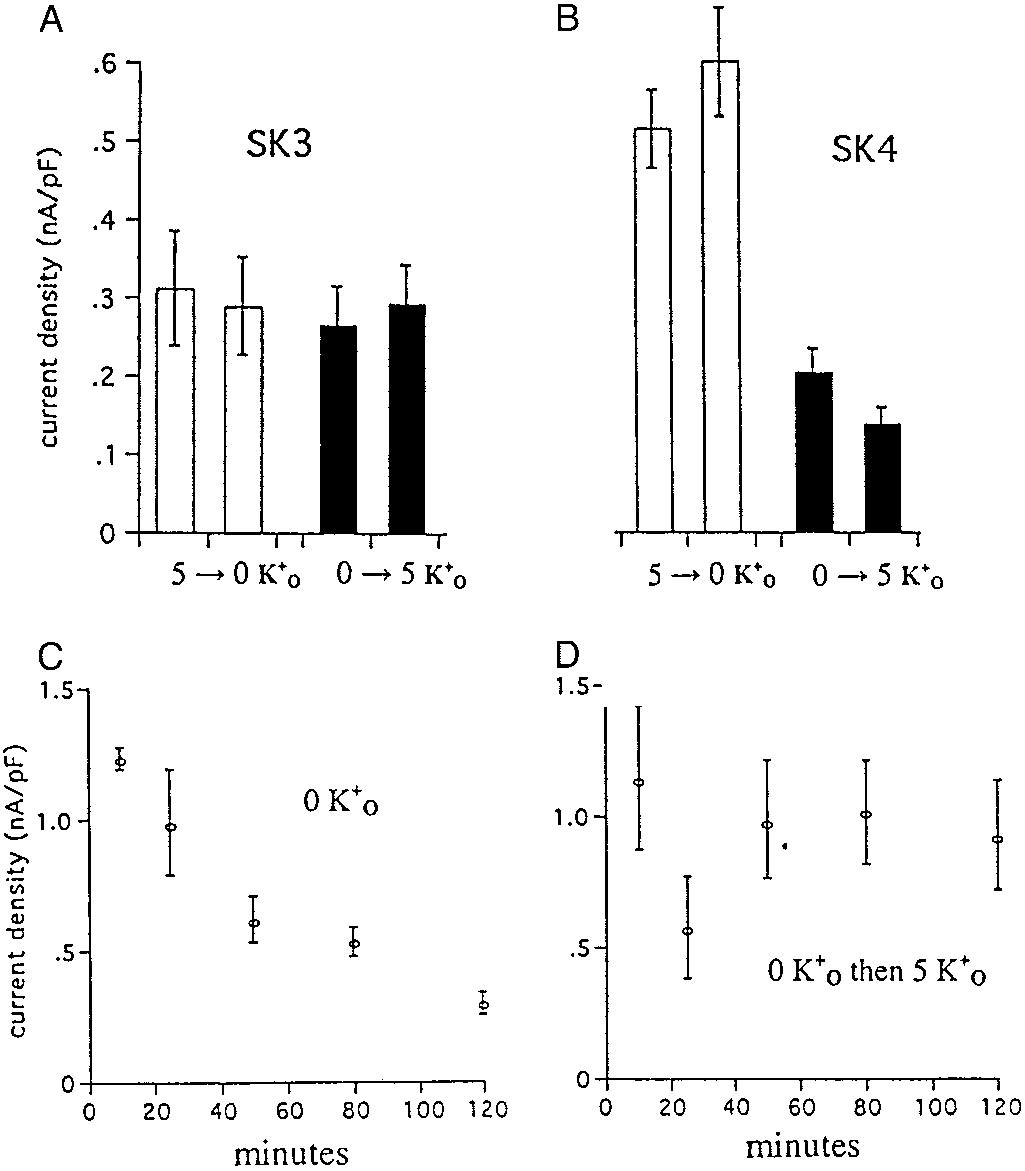
o on the activity of heterologously expressed
protein of the appropriate molecular weight is detected not only
SK3 and SK4 channels in CHO cells. Cells were washed twice in 0 mM Ko and
in the human erythroid progenitor cells but also in mature
then incubated with 100 M 1,2-bis(2-aminophenoxy)ethane-N,N,NЈ,NЈ-tetraacetate or -tetraacetic acid–acetoxymethyl ester (BAPTA-AM) in either 5
human erythrocyte membranes (21, 30). This signal was com-
mM Kϩ (open bars) or 0 mM Kϩ (dark bars) containing media for 3–5 h at room
pletely competed through preincubation of the antibody with the
temperature (see Methods). At the end of this period, cells were perfused with
antigenic peptide, demonstrating specificity of the interaction.
incubation solution before and during seal formation, breakthrough, and
Another indication of the specificity of the SK4 antibody is that
achievement of whole-cell recording mode. Current amplitude at 60 mV was
it showed no crossreactivity with brain, a tissue known to express
then measured for each cell first during continued perfusion with incubation
SK1, 2, and 3 but not the SK4 isoform.
solution and then after switching from 0 to 5 Ko or vice versa. The whole-cell
We next compared several functional characteristics of the
currents in expressed SK3 channels (A) were insensitive to the presence or
Gardos channel in human erythrocytes with the behavior of SK4
absence of Ko , whereas the currents in expressed SK4 channels (B) weremarkedly diminished by preincubation in 0 mM Kϩ. (C) The decrease in
expressed in transfected cells. The first modality to be examined
whole-cell currents that occurred during the first 2 h of incubation of SK4
expressing cells in 0 mM Ko . (D) SK4 cells can recover from incubation in 0 mM
It is known from previous work (9, 31) that for the Gardos
o by reexposure to 5 mM Ko over this time period. SK4 cells exposed for
longer periods of time (B, dark bars) to 0 mM Kϩ remain refractory to 5 mM Kϩ.
channel at the time that the cytoplasmic surface of the channel͞
Error bars represent Ϯ SEM, where n ϭ 9 –15 separate observations with SK3
(A) and 11–15 for SK4 (B); in C and D, n ϭ 3–12.
is introduced inside and, at least in human red
cell ghosts (9, 31), will not open even when Kϩo is subsequently
over this same time period. Thus, SK4 channels expressed in
added. We wished to test whether SK3 or SK4 channels heter-
CHO cells behave like the Gardos channel in intact human
ologously expressed in CHO cells displayed a similar depen-
erythrocytes and ghosts because they stay closed in the absence
dence on Kϩo. To this end, stably transfected CHO cells were
of Kϩ but differ from them in that the channels are able to open
preincubated for varying periods of time in the presence or
after the addition of Kϩ. The basis for this difference between
o . Whole-cell recordings were then made, with Cai
expressed SK4 channels and Gardos channels is not known, but
being introduced with a patch pipette, as described in Methods
it should be remembered that, because these patched CHO cells
and the Fig. 4 legend, with subsequent determination of the
are attached to the substrate, Kϩ trapped in the cell͞substrate
current density. As shown in Fig. 4A, the current measured in
interface may alter the overall response to Kϩ-free solutions.
cells expressing SK3 channels remained active and was insensi-
That the whole-cell currents do not fall to zero in Kϩo-free
solutions (Fig. 4B) may reflect the presence of this residual Ko .
cells expressing SK4 (Fig. 4B) were sensitive to the presence of
The second functional feature we explored was the temper-
Kϩo, displaying much less activity in cells preincubated without
ature sensitivity of the Gardos channel. Previous work (32) had
Kϩo. The cells studied in Fig. 4B were incubated in the absence
demonstrated that the open probability (Po) of Gardos channels,
o for 3–5 h and were refractory to the addition of Ko after
studied in inside-out patches of human erythrocytes, was re-
this time (data not shown). We then studied whether the
markably sensitive to temperature: the Po at 30°C was Ϸ0.6
expressed SK4 channel could be activated by Kϩo after shorter
falling to Ϸ0.1 at 37°C. We examined this effect of temperature
incubation times in its absence. Fig. 4C shows the time course of
on Gardos channel-mediated Kϩ (Rbϩ) flux in intact human
decay in SK4 activity during 2 h incubation in the absence of Kϩ
PHYSIOLOGY
erythrocytes as well as the response of the Po values of SK4
As shown in Fig. 4D, the cells can respond to the addition of Kϩo
channels expressed in CHO cells to changes in temperature.
PNAS ͉ June 10, 2003 ͉ vol. 100 ͉ no. 12 ͉ 7369
less than those obtained in patched red cell membranes (32),
because the Kϩ fluxes were faster at 27 than at 37°C.
Because of the above results, we were interested in evaluating the
temperature dependence of Ca2ϩ-activated Kϩ transport through
SK4 channels expressed in CHO cells as shown in Fig. 5. Fig. 5 A
and B show single-channel activity on two time scales recorded at
25 and 35°C. The results are summarized in the bar graphs (Fig. 5C),
where it is clear the Po value at 35°C is less than that at 25°C,
consistent with the previous intact red cell flux studies. The
observation that the differences in Po values are less than expected
based on previous results (32) may indicate that the membrane
environment surrounding SK4 channels expressed in CHO cells
exhibits substantial differences in the temperature-dependent lipid
phase transitions as compared with that present in red cells. The
differences may also be due to modulators such as calpromotin (24,
25), which is thought to be required for optimum Gardos activity.
We tested a purified sample of thioredoxin peroxidase that has been
shown to be identical to calpromotin (23). The results presented in
Fig. 5D indicate that it has no effect on the value of Po compared
with the controls (Fig. 5C). In addition, we prepared a hemolysate
from red cells according to established procedures (24, 25) that
should ensure it contained calpromotin; addition of this lysate to the
bathing medium in inside-out patches, as in Fig. 5D, was without
The sensitivity of hSK4 channels to changes in temperature. Results
of single-channel recordings from inside-out patches excised from CHO cells
Discussion
stably expressing hSK4 are presented. The bathing solution contained 1 Mfree Ca2ϩ with 130 mM Kϩ on both sides of the patch (see Methods). Record-
The main result of the work reported in this paper is that the
ings were performed at Ϫ80 mV first at 25°C and then, within 10 s, at 35°C. The
Gardos channel of human red blood cells is coded for by the
single-channel activity is shown in A and on an expanded time scale in B. The
human SK4 (i.e., KCNN4) gene, as described above in connec-
bars labeled 35°C in C and D represent the normalized values at 25°C (taken as
tion with Figs. 1–3. This is based first on analysis of RNA from
1.0) of the open probability (Po) of each channel from a given patch. The
human reticulocytes and cultured human erythroid progenitor
results presented in C represent control characteristics of SK4 channels,
cell, in which we found only the message for SK4 and second, on
whereas in D, thioredoxin peroxidase (see Methods) has been added to the
the use of an antibody directed against SK4. All of these
cytoplasmic bathing medium for reasons explained in the text. The error bars
preparations were characterized by ‘‘ profiling,’’ indicating that
are Ϯ SEM where n ϭ 6 in C and n ϭ 4 in D. Lumping the results of C and D
contamination from leukocytes and platelets was not detectable.
together, the mean difference between the values at 25 compared with 37°Cis 0.731 Ϯ 0.072 SEM, with P Ͻ 0.05.
Because SK4 is found in human leukocytes (3), this also means
that caution must be exercised in interpreting studies (e.g., ref.
7) where embryonic stem cells have been analyzed for SK and
The results of net Kϩ efflux experiments in human erythrocytes
other isoforms, given that these preparations are likely to be
are presented in Table 2. The details of the protocols varied for each
contaminated with nonerythroid forms (see, e.g., refs. 35 and
experiment (see figure legends and Methods). The primary aim was
36). Additional evidence is provided by Western analysis of
to pretreat cells in such a manner that when they were divided and
human progenitor cells and erythrocyte ghosts, where it is clearly
placed at the two different temperatures, the efflux characteristics
shown (Fig. 3) that the SK4 protein is present.
reflected differences in Gardos channel activity and not differences
Other support for the conclusion that the SK4 isoform is
contents, metabolic states, and possible interfering
responsible for the Gardos channel is found in the parallels in
membrane transport processes during their subsequent incubation.
function that are displayed in comparisons of human red cells
However, this at best is an assumption and is the reason for changing
and expressed SK4 channels. This is seen in the effects on
the protocols. When the cells contained ClϪ as the principal anion,
Ca2ϩ-activated Kϩ channels of preincubation of cells in the
i , regardless of protocol design, was the same at 27
presence and absence of Ko (Fig. 4) and in the decrease in Po
and 37°C (data not shown), similar to previous results (33). For
of channels when the temperature is raised from 25–27°C
instance, net Kϩ effluxes were also carried out as part of the same
to 35–37°C (Table 2 and Fig. 5). It should be emphasized,
experiment (B in Table 2) but where ClϪ replaced SCNϪ in the
however, that neither the parallel in the effects of Kϩo nor the
protocol. The comparable efflux rate constants (hrϪ1) at 27 and
temperature can be considered definitive, because their relative
37°C were 0.33 and 0.35, respectively. Because we thought that
effects fall short of expectations based on intact red cell͞ghost
the Kϩ efflux was rate limited by the membrane’s permeability to
results (9, 31) or determination of Po by patch analyses of red
ClϪ, i.e., PK͞PCl ϾϾ 1, we switched the principal anion to SCNϪ be-
cell membranes (32). On the other hand, previous observations
cause it has been shown that, under the conditions we were using,
by others, as referred to in the Introduction, have defined critical
PSCN ϾϾ PK (34). In the results shown in Table 2, the efflux of Kϩ,
biophysical and pharmacological properties that implicate
the SK4 isoform as being the Gardos channel in human red
than at 37°C. The variation in the flux values seen in the different
cells. Thus the single-channel conductance of heterologously
experiments is due primarily to the different protocols used. An
expressed SK4 channels as well as their electrical characteristics
important caveat in these experiments is that there may be heter-
and pharmacological profiles were essentially the same as
ogeneity in the response of the cell population to Gardos channel
that seen for Gardos channels in intact human red cells (7, 9, 10,
activity, because the extrapolated equilibrium end points varied and
13, 32). It is possible that the Gardos channel may have one or
were not necessarily the same for the two temperatures in most
more modulators, still to be defined, that are necessary for the
experiments (data not shown). Even so, the temperature depen-
hSK4 isoform to display fully the known characteristics of the
dence of Kϩ efflux (Table 2) in intact red cells appears to reflect
Gardos channel in human red blood cells. Candidates for such
changes in the Po of the Gardos channel that are consistent with but
modulators, in addition to the lipid environment, may be cal-
7370 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.1232342100
modulin (13, 30, 37, 38), redox systems (39, 40), and͞or protein
invaluable advice and help during the course of this work. In addition,
we thank Drs. G. Moss for the rSK3 construct and B. Kristensen for the
sample of thioredoxin peroxidase. This work was supported in part by
We thank Drs. M. Caplan, C. Canessa, V. Lew, K. Chandler, S.
National Institutes of Health Grant HL09906 (to J.F.H.) and by a grant
Basavappa, V. Rajendran, P. Bennekou, and P. Christophersen for
from the Wendy Will Case Cancer Fund (to A.W.).
1. Joiner, W. J., Wang, L.-Y., Tang, M. D. & Kaczmarek, L. K. (1997) Proc. Natl.
21. Joiner, W. J., Basavappa, S., Vidyasagar, S., Nehrke, K., Krishnan, S., Binder,
Acad. Sci. USA 94, 11013–11018.
H. J., Boulpaep, E. L. & Rajendran, V. M. (2003) Am. J. Physiol.,
2. Ishii, T. M., Silvia, C., Hirschberg, B., Bond, C. T., Adelman, J. P. & Maylie,
J. (1997) Proc. Natl. Acad. Sci. USA 94, 11651–11656.
22. Proverbio, F. & Hoffman, J. F. (1977) J. Gen. Physiol. 69, 605–632.
3. Logsdon, N. J., Kang, J., Togo, J. A., Christian, E. P. & Aiyar, J. (1997) J. Biol.
23. Kristensen, P., Rasmussen, D. E. & Kristensen, B. I. (1999) Biochem. Biophys.Chem. 272, 32723–32726. Res. Commun. 262, 127–131.
4. Ko¨hler, M., Hirschberg, B., Bond, C. T., Kinzie, J. M., Marrion, N. V., Maylie,
24. Plishker, G. A., White, P. H. & Cadman, E. D. (1986) Am. J. Physiol. 251,
J. & Adelman, J. P. (1996) Science 273, 1709–1714.
5. Warth, R., Hamm, K., Bleich, M., Kunzelmann, K., von Hahn, T., Schreiber,
25. Moore, R. B., Mankad, M. V., Shriver, S. K., Mankad, V. N. & Plishker, G. A.
R., Ullrich, E., Mengel, M., Trautmann, N., Kindle P., et al. (1999) Pflu¨gers
(1991) J. Biol. Chem. 266, 18964–18968. 438, 437–444.
6. Ghanshani, S., Coleman, M., Gustavsson, P., Wu, A. C., Gargus, J. J., Gutman,
26. Querales, D. B. P. (1999) Ph.D. thesis (Univ. of Cambridge, Cambridge, U.K.).
G. A., Dahl, N., Mahrenweiser, H. & Chandy, K. G. (1998) Genomics 51, 160–161.
27. Tiffert, T., Daw, N., Perdomo, D. & Lew, V. L. (2001) J. Lab. Clin. Med. 137,
7. Vandorpe, D. H., Shmukler, B. E., Jiang, L., Lim, B., Maylie, J., Adelman, J. P.,
de Franceschi, L., Cappellini, M. D., Brugnara, C. & Alper, S. L. (1998) J. Biol.
28. Heinz, A. & Hoffman, J. F. (1990) Proc. Natl. Acad. Sci. USA 87, 1998–2002. Chem. 273, 21542–21553.
29. Hoffman, J. F. (1962) J. Gen. Physiol. 45, 837–859.
8. Gardos, G. (1958) Biochim. Biophys. Acta 30, 653–654.
30. Joiner, W. J., Khanna, R., Schlichter, L. C. & Kaczmarek, L. K. (2001) J. Biol.
9. Grygorczyk, R., Schwarz, W. & Passow, H. (1984) Biophys. J. 45, 693–698. Chem. 276, 37980–37985.
10. Bennekou, P. & Christophersen, P. (2003) Red Cell Membrane Transport in
31. Heinz, A. & Passow, H. (1980) J. Membr. Biol. 57, 119–131. Health and Disease, eds. Bernhardt, I. & Ellory, J. C. (Springer, Berlin).
32. Grygorczyk, R. (1987) Biochim. Biophys. Acta 902, 159–168.
11. Castle, N. A. & Strong, P. N. (1986) FEBS Lett. 209, 117–121.
33. Simons, T. J. B. (1976) J. Physiol. 256, 209–225.
12. Alvarez, J., Montero, M. & Garcia-Sancho, J. (1992) J. Biol. Chem. 167,
34. Garcia-Sancho, J. & Lew, V. L. (1988) J. Physiol. 407, 523–539.
35. Guillemot, J.-C., Kruskal, B. A., Adra, C. N., Zhu, S., Ko, J.-L., Burch, P.,
13. Brugnara, C., Armsby, C. C., de Franceschi, L., Crest, M., Martin Eauclaire,
Nocka, K., Seetoo, K., Simons, E. & Lim, B. (1996) Blood 88, 2722–2731.
M.-F. & Alper, S. L. (1995) J. Membr. Biol. 147, 71–82.
36. Fibach, E., Manor, D., Oppenheim, A. & Rachmilewitz, E. A. (1989) Blood 73,
14. Shah, M. & Haylett, D. G. (2000) Br. J. Pharmacol. 129, 627–630.
15. Strobaek, D., Jorgensen, T. D., Christophersen, P., Ahring, P. K. & Olesen,
37. Hoffman, J. F., Yingst, D. R., Goldinger, J. M., Blum, R. M. & Knauf, P. A.
S.-P. (2000) Br. J. Pharmacol. 129, 991–999.
(1980) in Membrane Transport in Erythrocytes, eds. Lassen, U. V., Ussing, H. H.
16. Barfod, E. T., Moore, A. L. & Lidofsky, S. D. (2001) Am. J. Physiol. 280,
& Wieth, J. O. (Munksgaard, Copenhagen), pp. 178–195.
17. Jensen, B. S., Strobaek, D., Christophersen, P., Jorgensen, T. D., Hansen, C.,
38. Fanger, C. M., Ghanshani, S., Logsdon, N. J., Rauer, H., Kalman, K., Zhou,
Silahtaroglu, A., Olesen, S.-P. & Ahring, P. K. (1998) Am. J. Physiol. 275,
J., Bechingham, K., Chandy, K. G., Cahalan, M. D. & Aiyar, J. (1999) J. Biol.Chem. 274, 5746–5754.
18. Carignani, C., Roncarati, R., Rimini, R. & Terstappen, G. C. (2002) Brain Res.
39. Alvarez, J., Camaleno, J. M., Garcia-Sancho, J. & Herreros, B. (1986) Biochim.939, 11–18. Biophys. Acta 856, 408–411.
19. Stengelin, M. K. & Hoffman, J. F. (1997) Proc. Natl. Acad. Sci. USA 94,
40. Fuhrmann, G. F., Schwarz, W., Kersten, R. & Sdun, H. (1985) Biochim.Biophys. Acta 820, 223–234.
20. Hoffman, J. F., Wickrema, A., Potapova, O., Milanick, M. & Yingst, D. R.
41. Pellegrino, M. & Pellegrini, M. (1998) Pflu¨gers Arch. 436, 749–756.
(2002) Proc. Natl. Acad. Sci. USA 99, 14572–14577.
42. Andrews, D. A., Yang, L. & Low, P. S. (2002) Blood 100, 3392–3399. PHYSIOLOGY
PNAS ͉ June 10, 2003 ͉ vol. 100 ͉ no. 12 ͉ 7371
Standards der Raucherentwöhnung Konsensus der ÖGP Beilage zu Wien. Klin. Wochenschr. 117, Heft ■ (2005)Dieser Beitrag ist urheberrechtlich geschützt. Die dadurch begründeten Rechte, insbesondere die der Übersetzung, des Nachdruckes,der Entnahme von Abbildungen, der Funksendung, der Wiedergabe auf photomechanischem oder ähnlichem Wege und derSpeicherung in Datenverarbeitungsanlagen
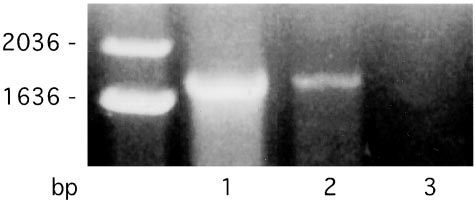
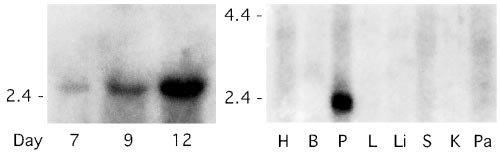 Table 2. The effect of temperature on the net efflux of K؉ from
Table 2. The effect of temperature on the net efflux of K؉ from
 Western blots showing that the protein for the Gardos channel
isoform, SK4, is present in cultured human erythroid progenitor cells and inghost membranes made from mature human red blood cells. The antibodywas prepared against an SK4-specific peptide and used as described in Meth-ods. The two positive controls are human parotid gland (P) and kidney (K) withthe negative control being brain (B). It is clear that a band of the appropriatemolecular weight is present in the human erythroid progenitor cells as theymature from days 7 to 13. It is also evident that the SK4 band is present inhuman red cell ghost membranes (RBC). The decrease in the blot intensity ofthe D13 band compared with D7 is primarily due to the decreased proteincontent (cell number) of cells loaded onto the gel. The slight variation in themolecular weights of SK4 bands seen in the progenitor cells, relative to theother bands, may be due to posttranslational modification or higher saltconcentration in the loading mixture. It should also be mentioned that, exceptin the parotid lane, there are higher molecular weight bands (not shown) thatin each case react with the antibody. Importantly, preincubation of theantibody with purified peptide that contains the antigenic epitope producesa complete loss of reactivity in all lanes except in brain, where it is muchreduced, and in ghosts, where it is only faintly present in the highest molecularweight bands (data not shown).
Western blots showing that the protein for the Gardos channel
isoform, SK4, is present in cultured human erythroid progenitor cells and inghost membranes made from mature human red blood cells. The antibodywas prepared against an SK4-specific peptide and used as described in Meth-ods. The two positive controls are human parotid gland (P) and kidney (K) withthe negative control being brain (B). It is clear that a band of the appropriatemolecular weight is present in the human erythroid progenitor cells as theymature from days 7 to 13. It is also evident that the SK4 band is present inhuman red cell ghost membranes (RBC). The decrease in the blot intensity ofthe D13 band compared with D7 is primarily due to the decreased proteincontent (cell number) of cells loaded onto the gel. The slight variation in themolecular weights of SK4 bands seen in the progenitor cells, relative to theother bands, may be due to posttranslational modification or higher saltconcentration in the loading mixture. It should also be mentioned that, exceptin the parotid lane, there are higher molecular weight bands (not shown) thatin each case react with the antibody. Importantly, preincubation of theantibody with purified peptide that contains the antigenic epitope producesa complete loss of reactivity in all lanes except in brain, where it is muchreduced, and in ghosts, where it is only faintly present in the highest molecularweight bands (data not shown).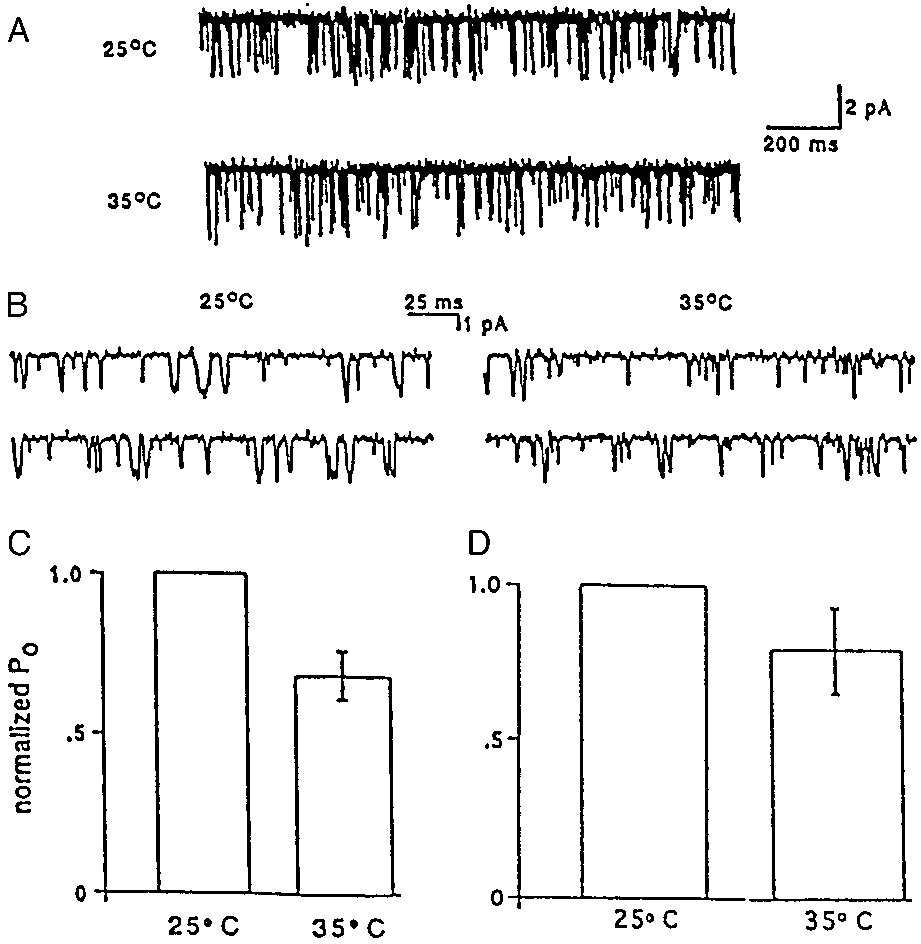 less than those obtained in patched red cell membranes (32),
because the Kϩ fluxes were faster at 27 than at 37°C.
less than those obtained in patched red cell membranes (32),
because the Kϩ fluxes were faster at 27 than at 37°C.