Botanical Journal of the Linnean Society, 2009, 161, 105–121. With 1 figure An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IIIboj_996105.121
1Recommended citation: APG III (2009). This paper was compiled by Birgitta Bremer, Kåre Bremer,Mark W. Chase, Michael F. Fay, James L. Reveal, Douglas E. Soltis, Pamela S. Soltis and Peter F. Stevens, who were equally responsible and listed here in alphabetical order only, with contributionsfrom Arne A. Anderberg, Michael J. Moore, Richard G. Olmstead, Paula J. Rudall, Kenneth J. Sytsma, David C. Tank, Kenneth Wurdack, Jenny Q.-Y. Xiang and Sue Zmarzty (in alphabeticalorder). Addresses: B. Bremer, The Bergius Foundation at the Royal Swedish Academy of Sciences, POBox 50017, SE-104 05 Stockholm, Sweden; K. Bremer, Vice Chancellor, Stockholm University, SE-10691 Stockholm, Sweden; M. W. Chase, M. F. Fay, Jodrell Laboratory, Royal Botanic Gardens, Kew,Richmond, Surrey, TW9 3DS, UK; J. L. Reveal, L.H. Bailey Hortorium, Department of Plant Biology,412 Mann Building, Cornell University, Ithaca, NY 14853-4301, USA; D. E. Soltis, Department ofBiology, University of Florida, Gainesville, Florida 32611–8525, USA; P. S. Soltis, Florida Museum ofNatural History, University of Florida, Gainesville, Florida, 32611–7800, USA; and P. F. Stevens,Department of Biology, University of Missouri-St. Louis and Missouri Botanical Garden, PO Box 299,St. Louis, Missouri 63166–0299, USAReceived 12 August 2009; accepted for publication 18 August 2009
A revised and updated classification for the families of flowering plants is provided. Many recent studies haveyielded increasingly detailed evidence for the positions of formerly unplaced families, resulting in a number ofnewly adopted orders, including Amborellales, Berberidopsidales, Bruniales, Buxales, Chloranthales, Escalloniales,Huerteales, Nymphaeales, Paracryphiales, Petrosaviales, Picramniales, Trochodendrales, Vitales and Zygophylla-les. A number of previously unplaced genera and families are included here in orders, greatly reducing the numberof unplaced taxa; these include Hydatellaceae (Nymphaeales), Haptanthaceae (Buxales), Peridiscaceae(Saxifragales), Huaceae (Oxalidales), Centroplacaceae and Rafflesiaceae (both Malpighiales), Aphloiaceae, Geisso-lomataceae and Strasburgeriaceae (all Crossosomatales), Picramniaceae (Picramniales), Dipentodontaceaeand Gerrardinaceae (both Huerteales), Cytinaceae (Malvales), Balanophoraceae (Santalales), Mitrastemonaceae(Ericales) and Boraginaceae (now at least known to be a member of lamiid clade). Newly segregated familiesfor genera previously understood to be in other APG-recognized families include Petermanniaceae (Liliales),Calophyllaceae (Malpighiales), Capparaceae and Cleomaceae (both Brassicales), Schoepfiaceae (Santalales),Anacampserotaceae, Limeaceae, Lophiocarpaceae, Montiaceae and Talinaceae (all Caryophyllales) and Linder-niaceae and Thomandersiaceae (both Lamiales). Use of bracketed families is abandoned because of its unpopu-larity, and in most cases the broader circumscriptions are retained; these include Amaryllidaceae, Asparagaceaceand Xanthorrheaceae (all Asparagales), Passifloraceae (Malpighiales), Primulaceae (Ericales) and several othersmaller families. Separate papers in this same volume deal with a new linear order for APG, subfamilial namesthat can be used for more accurate communication in Amaryllidaceae s.l., Asparagaceace s.l. and Xanthorrheaceaes.l. (all Asparagales) and a formal supraordinal classification for the flowering plants.
of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121.
ADDITIONAL KEYWORDS: angiosperm classification – phylogenetic classification – DNA phylogenetics –classification system – APG system.
*Corresponding author: Mark W. Chase. E-mail: [email protected]
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
along APG lines. They have all agreed to adoptAPG III as their standard and the linear order of
Since the previous APG classification (APG II, 2003),
Haston et al. (2009). In general, the broader cir-
a great deal more information about flowering plant
cumscriptions have been favoured and are adopted
relationships has emerged, and a more refined and
better-resolved classification is now possible. If a well-
2. Papers over the last few years have clarified the
supported hypothesis of monophyly is a necessary
positions of isolated families including Cerato-
prerequisite for a group to be named, it is not suffi-
phyllaceae, Chloranthaceae and Picramniaceae
cient. Not all clades need be named and, indeed, it
(Jansen et al., 2007; Moore et al., 2007; Wang
would barely be practicable to do this, so to decide
et al., 2009), and this has necessitated addition of
which clades should be named, additional criteria can
orders not previously recognized by APG.
be invoked. Backlund & Bremer (1998) provided a
3. A few genera/families, members of which had
useful discussion on the principles of rank-based phy-
either not been sequenced before or for which
logenetic classification that is applicable at all levels
chimaeric sequences were produced, were wrongly
apart from species and immediately above (see Albach
placed. Thus, families like Guamatelaceae have
et al., 2004; Entwisle & Weston, 2005; Pfeil & Crisp,
been added; Guamatela used to be placed in
2005, etc., for examples). We follow these principles
here. Backlund and Bremer’s main principle is that
Crossosomatales (Oh & Potter, 2006). Hydatel-
taxa that are recognized formally should be mono-
laceae have been moved from Poales to Nymphae-
phyletic. However, this does not indicate which par-
ticular clades should be named as families, orders etc.
4. There are a few cases where the general pattern of
It is helpful if (1) taxa formally recognized are
relationships has not changed much since APG II,
easily recognizable, (2) groups that are well estab-
but our appreciation of the pattern of variation
lished in the literature are preserved, (3) the size of
has. For example, this helps justify inclusion of
groups is taken into account (particularly small ones,
which should be combined with others whenever pos-
5. Finally, a few family circumscriptions suggested by
sible) and (4) nomenclatural changes are minimized
APG II did not reflect general usage, so we have
(Backlund & Bremer, 1998). Thus, on the one hand,
modified these, an example being the broadly
numerous small groups have little to recommend
circumscribed Brassicaceae, which are here split
them, as individually they summarize only a small
amount of information and tend to clog the memory,whereas groups that are too large may have few
In general, we have tried not to change the status
obvious shared traits that can be used to recognize
of families if they have long been recognized, unless
them. Asparagales and Lamiales in particular have a
there are other good reasons for combining them.
number of problems in this regard and include fami-
However, we have taken the opportunity to combine
lies that are still in a state of flux in this respect.
mono- or oligogeneric families. Most of the family-
Changes are being made to the classification
level changes mentioned below have resulted in the
adopted by APG II for several reasons, but bearing
families now recognized being relatively well charac-
the Backlund and Bremer guidelines in mind.
terized. However, we realize that it is almost impos-
1. In the earlier versions of the Angiosperm Phylog-
sible to achieve universal agreement for change – of
eny Group classification, alternative circumscrip-
any sort. Note that we do not see the APG classifica-
tions were permitted for some families. However,
tion as continuing to mutate for the indefinite future.
this seems unnecessary and more likely to cause
Given the amount of phylogenetic work that has
confusion than clarity. Here we indicate one of the
taken place in the last five years, the changes pro-
alternatives, a choice based in part on the circum-
posed here are modest. We hope the classification
scriptions adopted by textbooks (e.g. Judd et al.,
below will be found to be reasonable and, hence, will
2007), dictionaries (e.g. Mabberley, 2008) and the
not need much further change, although we do note
general literature. Note that, in preparation for
those few areas where there is particular phylo-
the third edition of The Plant Book, Mabberley
genetic uncertainty that may necessitate further
(2008: xi, 927, 929) consulted widely among tax-
revision of familial or ordinal limits. For further dis-
onomists about which alternative they preferred,
cussion on the variation in the groups discussed,
and, more recently, the issue of these alternative
potential apomorphies, etc., see the literature cited
circumscriptions was discussed by researchers
and Stevens (2001); particularly important recent
representing several European herbaria (e.g. K, E,
work includes Wang et al. (2009: rosids), Tank &
BM, P, G and the Dutch herbaria collectively) that
Donoghue (in press), Moore et al. (2008, in press: core
are in the process of reorganizing their collections
eudicots), Wurdack & Davis (2009: Malpighiales) and
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
Refulio-Rodriguez & Olmstead (2008, pers. comm.:
Austrobaileyaceae Croizat (1943), nom. cons.
For those researchers who wish to employ a formal,
$Schisandraceae Blume (1830), nom. cons. (includ-
higher-level classification for the land plants, Chase
& Reveal (2009) have proposed a system. It is based
Trimeniaceae L.S.Gibbs (1917), nom. cons.
on the recognition that most previous systems
Schisandraceae s.l. are well characterized.
have inflated the ranks of the angiosperms and the
other major groups of land plants. In this system,
Chloranthaceae R.Br. ex Sims (1820), nom. cons.
all embryophytes are a class, and, therefore, theangiosperms are recognized collectively as a single
Chloranthaceae are probably sister to magnoliids
subclass, Magnoliidae, which necessitated the use of
(Moore et al., 2007). Separate ordinal status is
superorders for the major angiosperm clades recog-
warranted because of their phylogenetic position;
they are also morphologically distinctive.
The general sequence of orders follows the left to
right sequence of the largely ladderized tree inHaston et al. (2009). Within orders, the sequence of
families is alphabetical. Symbols: *new family place-
ment; †newly recognized order for the APG system;
Canellaceae Mart. (1832), nom. cons.
§new family circumscription described in the text;
Winteraceae R.Br. ex Lindl. (1830), nom. cons.
$families that represent the broader circumscriptionof options available in APG II and favoured here,
$$families that were in square brackets in APG II,
Aristolochiaceae Juss. (1789), nom. cons.
the narrower circumscriptions favoured here. The list
Hydnoraceae C.Agardh (1821), nom. cons.
reflects a starting date for all flowering plant family
Lactoridaceae Engl. (1888), nom. cons.
names as 4 August 1789 (Jussieu, Genera plantarum).
Piperaceae Giseke (1792), nom. cons.
Full citations are available elsewhere (Reveal, 2008 –
Saururaceae F.Voigt (1811), nom. cons.
onward). A summarized phylogenetic tree represent-
The relationships of Hydnoraceae are unclear
ing the relationships among the major groups
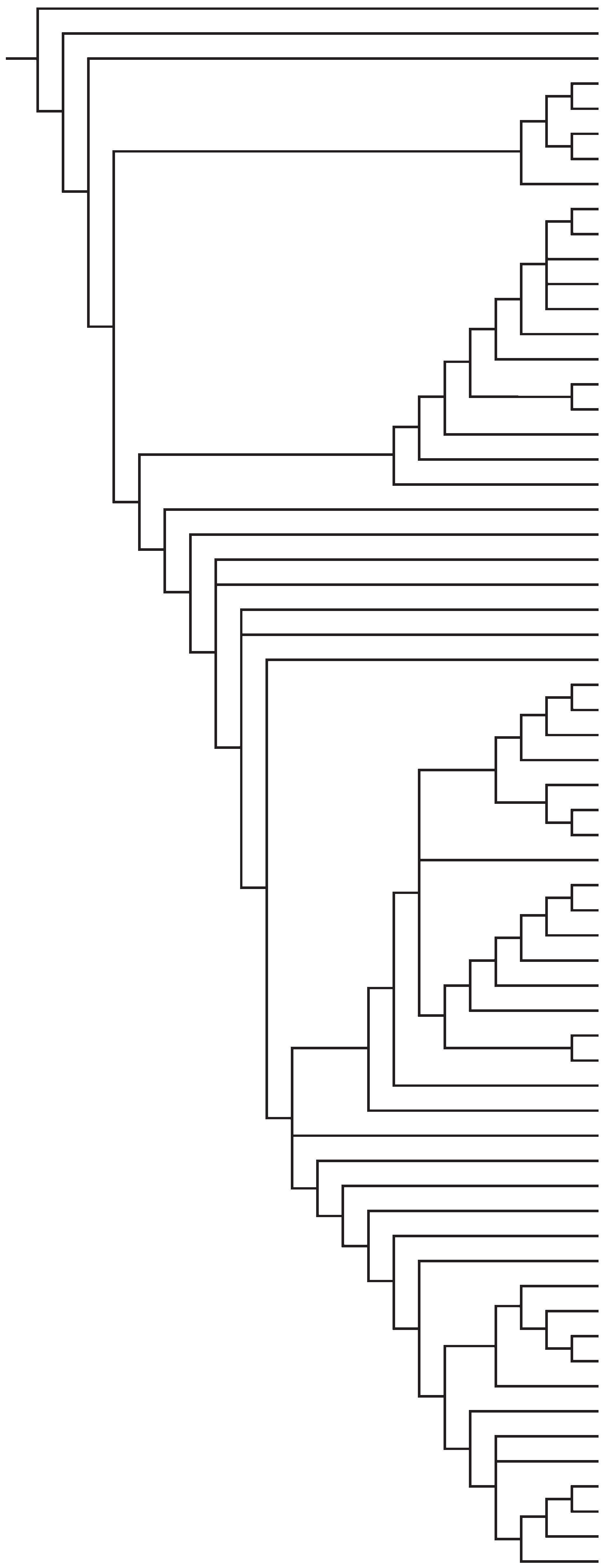
recognized here is presented in Figure 1.
Laurales Juss. ex Bercht. & J.Presl (1820)
Calycanthaceae Lindl. (1819), nom. cons. Gomortegaceae Reiche (1896), nom. cons.
†Amborellales Melikyan, A.V.Bobrov & Zaytzeva
Hernandiaceae Blume (1826), nom. cons.
Amborellaceae Pichon (1948), nom. cons.
Monimiaceae Juss. (1809), nom. cons.
The evidence that Amborellaceae are sister to all
other angiosperms is clear (e.g. Hansen et al., 2007;
Magnoliales Juss. ex Bercht. & J.Presl (1820)
Jansen et al., 2007; Moore et al., 2007). However, even
if they were sister to Nymphaeales (e.g. Goremykin,
Degeneriaceae I.W.Bailey & A.C.Sm. (1942), nom.
Viola & Hellwig, 2009), they should be kept separate
as their inclusion in Nymphaeales would result in a
Eupomatiaceae Orb. (1845), nom. cons.
Himantandraceae Diels (1917), nom. cons.
†Nymphaeales Salisb. ex Bercht. & J.Presl (1820)
Magnoliaceae Juss. (1789), nom. cons.
$$Cabombaceae Rich. ex A.Rich. (1822), nom. cons.
Myristicaceae R.Br. (1810), nom. cons.
*Hydatellaceae U.Hamann (1976)$$Nymphaeaceae Salisb. (1805), nom. cons.
There seems to be a general preference for keeping
Cabombaceae and Nymphaeaceae separate, although
both are small in terms of species. The two familiesare easily characterizable. The inclusion of Hydatel-
Alismatales R.Br. ex Bercht. & J.Presl (1820)
laceae (previously included in Poales) here was unex-
§Alismataceae Vent. (1799), nom. cons. (including
pected, but it is well supported morphologically and
embryologically (Saarela et al., 2007; Friedman, 2008;
Aponogetonaceae Planch. (1856), nom. cons.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
MagnolialesLauralesChloranthales†CommelinalesZingiberalesPoales
DasypogonaceaeAsparagalesLilialesPandanalesDioscorealesPetrosaviales†AlismatalesAcoralesCeratophyllales†RanunculalesSabiaceaeProteales
Buxales†Trochodendrales†GunneralesCucurbitalesFagalesRosalesFabalesCelastrales
Picramniales†CrossosomatalesMyrtalesGeranialesVitales†SaxifragalesDilleniaceaeBerberidopsidales†SantalalesCaryophyllalesCornalesEricales
SolanalesBoraginaceaeAquifolialesEscalloniales†AsteralesDipsacales
Figure 1. Interrelationships of the APG III orders and some families supported by jackknife/bootstrap percentages greater than 50 or Bayesian posterior probabilities greater than 0.95 in large-scale analyses of angiosperms. See text for literature supporting these relationships. Newly-recognized-for-APG orders are denoted (†). Some eudicot families not yet classified to order are not shown.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
Cymodoceaceae Vines (1895), nom. cons.
Hydrocharitaceae Juss. (1789), nom. cons.
Juncaginaceae Rich. (1808), nom. cons.
Posidoniaceae Vines (1895), nom. cons.
Ruppiaceae Horan. (1834), nom. cons.
Melanthiaceae Batsch ex Borkh. (1797), nom. cons.
Scheuchzeriaceae F.Rudolphi (1830), nom. cons.
*Petermanniaceae Hutch. (1934), nom. cons.
Philesiaceae Dumort. (1829), nom. cons.
Zosteraceae Dumort. (1829), nom. cons.
Ripogonaceae Conran & Clifford (1985)Smilacaceae Vent. (1799), nom. cons.
Alismataceae s.s. is lacking (e.g. Chen et al., 2004a, b)
Petermanniaceae are morphologically and phylo-
and the family does not have any apomorphies. When
genetically distinct. Luzuriagaceae, consisting of two
combined with Limnocharitaceae, a family that was
small genera with generalized lily floral morphology,
only relatively recently described (Cronquist, 1981),
are sister to Alstroemeriaceae and have the same
the enlarged family has several distinctive charac-
distinctive twisted petioles, so combination is in order
ters. It may be necessary to split off Maundia from
Juncaginaceae (Iles et al., 2009; S. von Mering & J. W.
Kadereit, pers. comm.), and Maundiaceae Nakai is
$Amaryllidaceae J.St.-Hil., nom. cons. (including
available. However, it might be better in this case to
Agapanthaceae F.Voigt, Alliaceae Borkh., nom.
create a larger single family for the larger clade. More
study is needed before another monogeneric family is
$Asparagaceae Juss. (1789), nom. cons. (including
Agavaceae Dumort., nom. cons., Aphyllanthaceae
Burnett, Hesperocallidaceae Traub, Hyacintha-
Petrosaviaceae Hutch. (1934), nom. cons.
ceae Batsch ex Borkh., Laxmanniaceae Bubani,Ruscaceae M.Roem., nom. cons., Themidaceae
The isolated position of Petrosaviaceae here is well
supported (e.g. Tamura et al., 2004; Chase et al.,
Blandfordiaceae R.Dahlgren & Clifford (1985)Boryaceae M.W.Chase, Rudall & Conran (1997)
Doryanthaceae R.Dahlgren & Clifford (1985)
Burmanniaceae Blume (1827), nom. cons.
Hypoxidaceae R.Br. (1814), nom. cons.
Dioscoreaceae R.Br. (1810), nom. cons.
Ixioliriaceae Nakai (1943)Lanariaceae R.Dahlgren & A.E.van Wyk (1988)
Thismiaceae J.Agardh (1858) may turn out to be in
Orchidaceae Juss. (1789), nom. cons.
a clade separate from Burmanniaceae and, similarly,
Tecophilaeaceae Leyb. (1862), nom. cons.
the morphologically distinctive Taccaceae Dumort.
$Xanthorrhoeaceae Dumort. (1829), nom. cons.
(1829), nom. cons., from Dioscoreaceae. Phylogenetic
(including Asphodelaceae Juss. and Hemerocalli-
relationships in Dioscoreales that support such
changes have been found by Merckx et al. (2006)
Xeronemataceae M.W.Chase, Rudall & M.F.Fay
and Merckx & Bidartondo (2008), and Merckx et al.
(2009) even suggest that Thismiaceae s.s. may beparaphyletic. Given the problems in understanding
An expanded Amaryllidaceae, including Agapan-
relationships of mycoheterotrophic groups, we have
thaceae and Alliaceae, are recognized here (Amaryl-
been conservative and not adopted any altered cir-
lidaceae was recently conserved over Alliaceae,
despite Alliaceae being the older name). Several char-acters support the combined group. Agapanthaceae, if
Pandanales R.Br. ex Bercht. & J.Presl (1820)
kept separate, are weakly characterized; the family is
Cyclanthaceae Poit. ex A.Rich. (1824), nom. cons.
Pandanaceae R.Br. (1810), nom. cons.
The area around Asparagaceae is difficult from the
Stemonaceae Caruel (1878), nom. cons.
standpoint of circumscription. Although Asparagaceae
Triuridaceae Gardner (1843), nom. cons. s.l. are heterogeneous and poorly characterized,
Velloziaceae J.Agardh (1858), nom. cons.
Asparagaceae s.s., Agavaceae, Laxmanniaceae, Rus-
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
caceae and even Hyacinthaceae have few if any dis-
Restionaceae R.Br. (1810), nom. cons.
tinctive features. Agavaceae and Ruscaceae sensu APG
Thurniaceae Engl. (1907), nom. cons.
II (2003) are heterogeneous and have been divided into
§Typhaceae Juss. (1789), nom. cons. (including
several families in the past, further confusing the
issue. This solution at least keeps the number of
Xyridaceae C.Agardh (1823), nom. cons.
difficult-to-recognize families to a minimum; Amaryl-
Sparganiaceae are included in Typhaceae; the two
lidaceae s.l. and Asparagaceae s.l. are easily differen-
families are monogeneric, occupy similar habitats and
tiated by umbels with a pair of enclosing bracts vs.
share a number of features. That they were treated
racemes or rarely umbels, but if umbels are present
separately in APG II was a mistake (M. W. Chase,
they contain three or more bracts (e.g. Brodiaea, Milla
pers. comm.). They have in the past been combined;
and relatives, Themidaceae) and lack the enclosing
Mabberley (2008) suggested that their combination
pair of bracts. Xanthorrhoeaceae s.l. have been
maintained for similar reasons to Asparagaceaes.l. Xanthorrhoeaceae s.s. are monogeneric, and
Asphodelaceae are impossible to distinguish from
genera such as Anthericum in Asparagaceae s.l.
Asparagaceae and Xanthorrhoeaceae sensu APG III is
Marantaceae R.Br. (1814), nom. cons.
proposed in Chase, Reveal & Fay (2009). This will
allow researchers to use a subfamily name where
Strelitziaceae Hutch. (1934), nom. cons.
previously they would have used one of the APG II
Zingiberaceae Martinov (1820), nom. cons.
Ceratophyllaceae Gray (1822), nom. cons.
Relationships of Dasypogonaceae remain unclear.
The molecular evidence that Ceratophyllaceae are
They are not particularly distinctive morphologically
sister to eudicots is becoming clearer (Jansen et al.,
and so do not warrant immediate ordinal status,
2007; Moore et al., 2007, but cf. Goremykin et al.,
although Dasypogonales Doweld is available. They
2009). In this and all other relationships that have
could probably be combined with one of the other
been suggested for Ceratophyllaceae, including sister
commelinid orders should they fall as sister groups.
to the monocots or Chloranthaceae (Endress & Doyle,2009), they are morphologically divergent from their
Arecaceae Bercht. & J.Presl (1820), nom. cons.
Commelinales Mirb. ex Bercht. & J.Presl (1820)
Commelinaceae Mirb. (1804), nom. cons. Haemodoraceae R.Br. (1810), nom. cons.
Ranunculales Juss. ex Bercht. & J.Presl (1820)
Berberidaceae Juss. (1789), nom. cons.
Philydraceae Link (1821), nom. cons.
Pontederiaceae Kunth (1816), nom. cons.
Eupteleaceae K.Wilh. (1910), nom. cons.
Lardizabalaceae R.Br. (1821), nom. cons.
Anarthriaceae D.F.Cutler & Airy Shaw (1965)
Menispermaceae Juss. (1789), nom. cons.
Bromeliaceae Juss. (1789), nom. cons.
$Papaveraceae Juss. (1789), nom. cons. (including
Centrolepidaceae Endl. (1836), nom. cons.
Fumariaceae Marquis, nom. cons., Pteridophyl-
Ecdeiocoleaceae D.F.Cutler & Airy Shaw (1965)
Ranunculaceae Juss. (1789), nom. cons.
Eriocaulaceae Martinov (1820), nom. cons. Flagellariaceae Dumort. (1829), nom. cons.
We adopt broad limits for Circaeasteraceae and
Joinvilleaceae Toml. & A.C.Sm. (1970)
Papaveraceae, as this is commonly done (Judd et al.,
2007; Mabberley, 2008), and the two families are
well characterized in their broader circumscriptions.
The two families into which Circaeasteraceae have
Rapateaceae Dumort. (1829), nom. cons.
been divided (Circaeasteraceae and Kingdoniaceae)
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
are both monogeneric; they are herbaceous and
This family has no stable position as yet (Moore
their leaves have the same distinctive dichotomous
et al., in press). The ordinal name, Dilleniales DC. ex
Bercht. & J.Presl, is available.
Saxifragales Bercht. & J.Presl (1820)
Altingiaceae Horan. (1841), nom. cons.
Although Moore et al. (2008) placed Sabiaceae as
Cercidiphyllaceae Engl. (1907), nom. cons.
However, if further work confirms this position,
Crassulaceae J.St.-Hil. (1805), nom. cons.
Sabiaceae will be included in a broadened circum-
Daphniphyllaceae Müll.-Arg. (1869), nom. cons.
scription of Proteales; the two have features in
Grossulariaceae DC. (1805), nom. cons.
common. Sabiaceae remain poorly known.
$$Haloragaceae R.Br. (1814), nom. cons.
Proteales Juss. ex Bercht. & J.Presl (1820)
Hamamelidaceae R.Br. (1818), nom. cons.
Nelumbonaceae A.Rich. (1827), nom. cons.
$Iteaceae J.Agardh (1858), nom. cons. (including
$$Platanaceae T.Lestib. (1826), nom. cons.
$$Proteaceae Juss. (1789), nom. cons.
Paeoniaceae Raf. (1815), nom. cons. $$Penthoraceae Rydb. ex Britt. (1901), nom. cons.
Platanaceae, although monogeneric, are morpho-
logically distinct from Proteaceae, and the two have
(including Medusandraceae Brenan, nom. cons.,
never been combined previously; members of the
broader family would have few features in common.
Saxifragaceae Juss. (1789), nom. cons.
†Trochodendrales Takht. ex Cronquist (1981)
The limits of Iteaceae are broadened because the
combined clade is well characterized and Pteroste-
monaceae are monogeneric. The limits of Halor-
A separate order for this morphologically distinct
agaceae are drawn narrowly as the inclusion of
clade is warranted; the two monospecific genera in
Penthoraceae and Tetracarpaeaceae would result in
a family with no obvious characters and totally
dendron, have much in common.
novel limits. The three families are individually tol-erably well characterized. Recent molecular analyses
$Buxaceae Dumort. (1822), nom. cons. (including
within Saxifragales, as sister to all other members
of this clade (Soltis et al., 2007a; Jian et al., 2008),
rather than in Malpighiales as previously proposed.
The limits of Buxaceae are expanded. The monoge-
Peridiscaceae continue to be expanded (Davis &
neric Didymelaceae have the same distinctive pollen
Chase, 2004; Wurdack & Davis, 2009), but all
and chemistry as at least part of Buxaceae, although
members of the expanded family have similar dis-
there is currently no evidence for the paraphyly of the
latter. Some morphological features suggest that
Cynomoriaceae are another family of holoparasitic
Haptanthaceae are best placed here, but they are
angiosperms that have been difficult to place. Some
distinct from all other angiosperms (Doust & Stevens,
molecular analyses had placed them in Santalales
2005). An order for the two families is warranted.
(Jian et al., 2008), although with little support.
Note that relationships of Trochodendrales and
However, Barkman et al. (2007) found no support for
Buxales remain unclear, although they are certainly
a position in that order or anywhere else. Neverthe-
to be placed in this part of the tree.
less, Nickrent (2002) and Nickrent et al. (2005)suggested that Cynomoriaceae should be placed inSaxifragales, but the evidence for placing them hereversus in Santalales is not strong. Confounding the
placement of this taxon is evidence for horizontal
gene transfer involving its host for some mitochon-
$$Gunneraceae Meisn. (1842), nom. cons.
drial genes (Barkman et al., 2007). Zhang, Li & Li
$$Myrothamnaceae Nied. (1891), nom. cons.
(2009) analyzed sequences from the plastid invertedrepeat and found that Cynomorium fell as sister
The two families share no important features and
to Rosaceae (Rosales) with high bootstrap support
so are kept separate, although both are monogeneric.
(99%). Due to these discordant results, we do not
Dilleniaceae Salisb. (1807), nom. cons.
assign Cynomoriaceae to an order here.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
Balanopaceae Benth. & Hook.f. (1880), nom. cons. Bonnetiaceae L.Beauvis. ex Nakai (1948)
†Vitales Juss. ex Bercht. & J.Presl (1820)
Caryocaraceae Voigt (1845), nom. cons.
Vitaceae remain isolated and ordinal status is appro-
*Centroplacaceae Doweld & Reveal (2005)
priate. They are sister to the fabids + malvids (rosid
$$Chrysobalanaceae R.Br. (1818), nom. cons.
I + II) clade in most recent analyses, albeit without
§Clusiaceae Lindl. (1836), nom. cons.
strong support (reviewed in Wang et al., 2009).
Ctenolophonaceae Exell & Mendonça (1951)$$Dichapetalaceae Baill. (1886), nom. cons.
Elatinaceae Dumort. (1829), nom. cons. $$§Erythroxylaceae
$$Krameriaceae Dumort. (1829), nom. cons.
Euphorbiaceae Juss. (1789), nom. cons.
$$Zygophyllaceae R.Br. (1814), nom. cons.
Although Krameriaceae are monogeneric, they are
Humiriaceae A.Juss. (1829), nom. cons.
Zygophyllaceae; the two are sister taxa. Combin-
Hypericaceae Juss. (1789), nom. cons.
ing the two would simply make a heterogeneous
Irvingiaceae Exell & Mendonça (1951), nom. cons.
Zygophyllaceae still more so. An order is needed for
Ixonanthaceae Planch. ex Miq. (1858), nom. cons.
this family pair as it is placed with strong support as
Lacistemataceae Mart. (1826), nom. cons.
sister to a clade containing more than two fabid
Linaceae DC. ex Perleb (1818), nom. cons.
orders in the analysis by Wang et al. (2009).
Lophopyxidaceae H.Pfeiff. (1951)Malpighiaceae Juss. (1789), nom. cons.
$Ochnaceae DC. (1811), nom. cons. (including
$Celastraceae R.Br. (1814), nom. cons. (including
Lepuropetalaceae Nakai, Parnassiaceae Marti-
Pandaceae Engl. & Gilg (1912–1913), nom. cons.
Lepidobotryaceae J.Léonard (1950), nom. cons.
$Passifloraceae Juss. ex Roussel (1806), nom. cons.
The limits of Celastraceae are broadened because
[including Malesherbiaceae D.Don, nom. cons.,
the three small families included show every sign
Turneraceae Kunth ex DC. (1828), nom. cons.]
of making Celastraceae paraphyletic if excluded
Phyllanthaceae Martinov (1820), nom. cons.
(Zhang & Simmons, 2006). The dismemberment of
Picrodendraceae Small (1917), nom. cons.
Celastraceae needed to maintain the families, of
Podostemaceae Rich. ex Kunth (1816), nom. cons.
which only Parnassiaceae are well known, would be
extensive and yield poorly characterized families, and
*Rafflesiaceae Dumort. (1829), nom. cons.
Celastraceae s.l. are better characterized than Celas-
$$Rhizophoraceae Pers. (1807), nom. cons.
traceae, excluding Parnassiaceae (for morphology, see
$$Trigoniaceae A.Juss. (1849), nom. cons. Violaceae Batsch (1802), nom. cons.
Brunelliaceae Engl. (1897), nom. cons.
The holoparasitic Rafflesiaceae are best assigned
Cephalotaceae Dumort. (1829), nom. cons.
to Malpighiales, perhaps making Euphorbiaceae s.s.
Connaraceae R.Br. (1818), nom. cons.
paraphyletic (e.g. Davis & Wurdack, 2004; Davis
Cunoniaceae R.Br. (1814), nom. cons. et al., 2007); the recognition of Peraceae Klotzsch
Elaeocarpaceae Juss. ex DC. (1816), nom. cons.
(1859) would be needed to maintain monophyly of
Euphorbiaceae. However, pending further studies,
Oxalidaceae R.Br. (1818), nom. cons.
Peraceae are not recognized here. Limits of clades inthe Bonnetiaceae–Podostemaceae area are becoming
Huaceae are tentatively included in Oxalidales
clearer (Wurdack & Davis, 2009), and this necessi-
because a number of recent studies (e.g. Wurdack &
tates the removal of Calophyllaceae from Clusiaceae.
Davis, 2009) have indicated that they are sister to
The alternatives would be a family that included both
Oxalidales as recognized in previous versions of APG.
of these families and Bonnetiaceae, Hypericaceae and
This is not a well-characterized clade, and it remains
Podostemaceae or one that included the last two
families plus Calophyllaceae; in both cases Hyperi-
Malpighiales Juss. ex Bercht. & J.Presl (1820)
caceae would be the correct name. The four families
Achariaceae Harms (1897), nom. cons.
in the area of Chrysobalanaceae, Dichapetalaceae,
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
Euphroniaceae and Trigoniaceae are kept separate
similar, even although Rhoipteleaceae have superior
as, although clearly related, a broadly drawn Chryso-
and Juglandaceae inferior ovaries; ovary position is
balanaceae would be heterogeneous (see Matthews &
variable in many families, and in other such cases,
Endress, 2008, for the morphology of this group).
e.g. Ebenaceae (including Lissocarpaceae), we have
Bhesa (formerly of Celastraceae) and Centroplacus
(formerly of Euphorbiaceae) form an isolated clade
(Davis et al., 2005; Wurdack & Davis, 2009) that has
Barbeyaceae Rendle (1916), nom. cons.
distinguishing features; recognition of a bigeneric
Cannabaceae Martinov (1820), nom. cons.
broadly drawn, i.e. including Samydaceae Vent. and
Elaeagnaceae Juss. (1789), nom. cons.
Scyphostegiaceae Hutch. Although the combined
Moraceae Gaudich. (1835), nom. cons.
clade is only moderately distinct morphologically and
less so phylogenetically (Chase et al., 2002), recogni-
tion of more families in this area is premature
pending a more detailed sampling of the genera. Trichostephanus Gilg, unplaced previously, shareswood anatomy, disc lobing and seed structure withSamydeae Vent. (= Salicaceae), and unpublished DNA
data support this placement (M. Alford, pers. comm.).
Geraniales Juss. ex Bercht. & J.Presl (1820)
$Geraniaceae Juss. (1789), nom. cons. (including
sister taxon, Erythroxylaceae, although Aneulophus,
of Erythroxylaceae, is to a certain extent morphologi-
$Melianthaceae Horan. (1834), nom. cons. (includ-
cally intermediate; the two families have hitherto not
been combined. Passifloraceae and Ochnaceae are
§Vivianiaceae Klotzsch, nom. cons. prop. (including
broadly delimited here yet remain readily character-
izable; relationships between the component cladeswithin the two families are uncertain, and both
Geraniales are a heterogeneous and poorly known
Medusagynaceae (Ochnaceae s.l.) and Malesherbi-
order. The inclusion of the monogeneric Hypseochari-
aceae (Passifloraceae s.l.) are monogeneric.
taceae in Geraniaceae, monogeneric Francoaceaein Melianthaceae and bigeneric Ledocarpaceae in
Cucurbitales Juss. ex Bercht. & J.Presl (1820)
Vivianiaceae leaves these expanded families with a
Begoniaceae C.Agardh (1824), nom. cons. Coriariaceae DC. (1824), nom. cons.
Myrtales Juss. ex Bercht. & J.Presl (1820)
Corynocarpaceae Engl. (1897), nom. cons.
Cucurbitaceae Juss. (1789), nom. cons.
Combretaceae R.Br. (1810), nom. cons.
Datiscaceae Dumort. (1829), nom. cons.
Crypteroniaceae A.DC. (1868), nom. cons.
Lythraceae J.St.-Hil. (1805), nom. cons. $Melastomataceae Juss. (1789), nom. cons. (includ-
§Myrtaceae Juss. (1789), nom. cons. (including
Polygalaceae Hoffmanns. & Link (1809), nom. cons.
Heteropyxidaceae Engl. & Gilg, nom. cons.,
§Penaeaceae Sweet ex Guill. (1828), nom. cons.
(including Oliniaceae Arn., nom. cons., Rhyn-
Casuarinaceae R.Br. (1814), nom. cons.
chocalycaceae L.A.S.Johnson & B.G.Briggs)
Vochysiaceae A.St.-Hil. (1820), nom. cons.
§Juglandaceae DC. ex Perleb (1818), nom. cons.
In Melastomataceae, and still more in Myrtaceae,
[including Rhoipteleaceae Hand.-Mazz. (1932),
common usage is for broadened family circumscrip-
tions. Both Heteropyxidaceae and Psiloxylaceae are
Myricaceae A.Rich. ex Kunth (1817), nom. cons.
small families and when included in Myrtaceae s.l.
that family remains characterized by possession of
Ticodendraceae Gómez-Laur. & L.D.Gómez (1991)
pellucid glands containing ethereal oils.
Juglandaceae are expanded to include their sister
clade, the monogeneric Rhoipteleaceae. The two look
Penaeaceae, Oliniaceae, Alzataeaceae and Rhyn-
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
chocalycaceae is clear; Van Beusekom-Osinga & van
Beusekom (1975) included the last two families in the
$Akaniaceae Stapf (1912), nom. cons. (including
Bretschneideraceae Engl. & Gilg, nom. cons.)
although they show variation in floral morphology
Bataceae Mart. ex Perleb (1838), nom. cons.
and embryo sac, in particular. Some combination is in
§Brassicaceae Burnett (1835), nom. cons.
order, and Penaeaceae have been expanded to include
*Capparaceae Juss. (1789), nom. cons.
Rhynchocalycaceae and Oliniaceae; Penaeaceae s.l.
Caricaceae Dumort. (1829), nom. cons.
Gyrostemonaceae A.Juss. (1845), nom. cons.
Crossosomataceae Engl. (1897), nom. cons.
Koeberliniaceae Engl. (1895), nom. cons.
Limnanthaceae R.Br. (1833), nom. cons.
*Guamatelaceae S.Oh & D.Potter (2006)
Moringaceae Martinov (1820), nom. cons.
Stachyuraceae J.Agardh (1858), nom. cons.
Pentadiplandraceae Hutch. & Dalziel (1928)
Staphyleaceae Martinov (1820), nom. cons.
Resedaceae Martinov (1820), nom. cons.
Salvadoraceae Lindl. (1836), nom. cons.
(including Ixerbaceae Griseb. ex Doweld & Reveal)
Setchellanthaceae Iltis (1999)Tovariaceae Pax (1891), nom. cons.
The addition of several families to Crossosomatales
Tropaeolaceae Juss. ex DC. (1824), nom. cons.
is well justified (e.g. Sosa & Chase, 2003; Soltis et al.,2007b; Wang et al., 2009), although monogeneric
Inclusion of monogeneric Bretschneideraceae into
Guamatelaceae are a somewhat surprising addition,
the monogeneric Akaniaceae is justified by the
morphological similarities of the two, which are
Rosaceae (Oh & Potter, 2006). For the most part,
sister taxa. Although a broad circumscription of
relationships among families included in Crossoso-
Brassicaceae was recognized in APG (1998) and APG
matales have not been suggested before. The order is
II (2003), the consensus prefers the recognition of
heterogeneous, although the families are all small.
three families, all of which can be characterized,
Nevertheless, Matthews & Endress (2005b, 2006)
albeit Capparaceae only rather poorly so. The final
found a number of floral features to be at least
phylogenetic positions, and hence taxonomic disposi-
common in the order. The sister taxa Strasburgeri-
tion, of some genera, particularly those previously
aceae and Ixerbaceae are two monogeneric families
included in Capparaceae–Stixeae, remain uncertain
that agree in several characters, including base chro-
(Hall, Sytsma & Iltis, 2002; Hall, Iltis & Sytsma,
mosome number and stamen and gynoecial morphol-
2004). Nonetheless, the name Stixaceae Doweld
ogy etc.; combination is in order. As the order is now
(2008) is available if it is required.
defined, Staphyleales Mart. (1835) is an older name
Malvales Juss. ex Bercht. & J.Presl (1820)
for Crossosomatales Takht. ex Reveal (1993). It need
$Bixaceae Kunth (1822), nom. cons. (including
Cochlospermaceae Planch., nom. cons., Diegoden-
*Picramniaceae Fernando & Quinn (1995)
Cistaceae Juss. (1789), nom. cons. *Cytinaceae A.Rich. (1824)
Picramniaceae were previously unplaced rosids, but
Dipterocarpaceae Blume (1825), nom. cons.
there is now strong support for a position in the rosid
II/malvid clade (Wang et al., 2009).
Muntingiaceae C.Bayer, M.W.Chase & M.F.Fay
*Dipentodontaceae Merr. (1941), nom. cons.
Neuradaceae Kostel. (1835), nom. cons.
Sarcolaenaceae Caruel (1881), nom. cons.
Sphaerosepalaceae Tiegh. ex Bullock (1959)Thymelaeaceae Juss. (1789), nom. cons.
This assemblage of three small families is well
supported, and recognition of Huerteales is appro-
A broad circumscription for Bixaceae is adopted; the
priate given their position. Within Huerteales, the
three families included are all small, and the combined
recently described Gerrardinaceae (Gerrardina was
family can be characterized morphologically. The para-
previously placed in Flacourtiaceae) are sister to the
sitic Cytinaceae (including Bdallophyton Eichl.) find
rest, and Dipentodontaceae, although monogeneric
their resting place here (Nickrent, 2007). The novel
like Gerrardinaceae, are distinctive (see Worberg
dismemberment of Malvaceae by Cheek (2006), see
also Cheek in Heywood et al., 2007) is not followed
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
here; the families are difficult to distinguish, and two
are not proposed here. Santalaceae are kept with
are new (Brownlowiaceae, Durionaceae, although the
their previous circumscription. That they can be
first is a later name for Sparmanniaceae J.Agardh as
divided into clades (Der & Nickrent, 2008), one of
defined by Cheek). The close relationship of the four
which is the morphologically distinct Viscaceae, is of
families that make up Malvaceae s.l. here has been
itself insufficient reason for their dismemberment
recognized since at least the time of Robert Brown.
(see Introduction, also Dipsacales below). Balanopho-
Details of relationships in the area of Cistaceae–
raceae are to be included in Santalales (Nickrent, Der
& Anderson, 2005; Barkman et al., 2007), and there is
and these families may need to be combined (Kubitzki
some evidence that Cynomoriaceae might also belong
& Chase, 2002; Ducousso et al., 2004); Cistaceae has
here (see comments under Saxifragales, above).
priority if these are all combined as a single family.
Caryophyllales Juss. ex Bercht. & J.Presl (1820)
Sapindales Juss. ex Bercht. & J.Presl (1820)
Achatocarpaceae Heimerl (1934), nom. cons.
Anacardiaceae R.Br. (1818), nom. cons.
Aizoaceae Martinov (1820), nom. cons.
Amaranthaceae Juss. (1789), nom. cons.
Burseraceae Kunth (1824), nom. cons.
*Anacampserotaceae Eggli & Nyffeler (2010, in
Ancistrocladaceae Planch. ex Walp. (1851), nom.
$Nitrariaceae Lindl. (1835), nom. cons. (including
Peganaceae Tiegh. ex Takht., Tetradiclidaceae
Asteropeiaceae Takht. ex Reveal & Hoogland (1990)
Sapindaceae Juss. (1789), nom. cons.
Simaroubaceae DC. (1811), nom. cons.
Caryophyllaceae Juss. (1789), nom. cons. §Didiereaceae Radlk. (1896), nom. cons.
We circumscribe Nitrariaceae broadly. The four
Dioncophyllaceae Airy Shaw (1952), nom. cons.
Droseraceae Salisb. (1808), nom. cons.
although their basic morphology, anatomy and chem-
Drosophyllaceae Chrtek, Slavíková & Studnicˇka
Frankeniaceae Desv. (1817), nom. cons.
Aextoxicaceae Engl. & Gilg (1920), nom. cons.
Halophytaceae A.Soriano (1984)*Limeaceae Shipunov ex Reveal (2005)
The morphologically distinct Aextoxicaceae and
*Lophiocarpaceae Doweld & Reveal (2008)
Berberidopsidaceae are strongly supported as sister
§Molluginaceae Bartl. (1825), nom. cons.
taxa, and recent work (Moore et al., in press) placed
them with strong support as sister to (Santalales
Nepenthaceae Dumort. (1829), nom. cons.
(Caryophyllales + asterids)); thus, ordinal status is
Nyctaginaceae Juss. (1789), nom. cons.
Santalales R.Br. ex Bercht. & J.Presl (1820)
Phytolaccaceae R.Br. (1818), nom. cons.
*Balanophoraceae Rich. (1822), nom. cons.
Plumbaginaceae Juss. (1789), nom. cons.
Loranthaceae Juss. (1808), nom. cons.
Polygonaceae Juss. (1789), nom. cons.
Misodendraceae J.Agardh (1858), nom. cons.
§Portulacaceae Juss. (1789), nom. cons.
Santalaceae R.Br. (1810), nom. cons.
Opiliaceae Valeton (1886), nom. cons.
Stegnospermataceae Nakai (1942)*Talinaceae Doweld (2001)
The genera included in Schoepfiaceae used to be
Tamaricaceae Link (1821), nom. cons.
included in Olacaceae s.l., but they are exclusivelyrelated. They are well supported as being in a clade
The recognition of a number of new but small
with Misodendraceae, but that family is morphologi-
families is necessitated by recent phylogenetic work on
cally so distinct that combination of the two families
core Caryophyllales. Anacampserotaceae, Portula-
is inappropriate. The paraphyletic Olacaceae are
caceae s.s., Montiaceae and Talinaceae are all clades
being resolved into a number of clades (Malécot &
near Cactaceae that are for the most part well
Nickrent, 2008), but relationships between these
supported as distinct (e.g. Applequist & Wallace, 2001;
clades are uncertain and so new families/family limits
Nyffeler, 2007; Nyffeler & Eggli, in press; Brockington
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121 et al., in press). Expansion of Cactaceae to include all
Roridulaceae Martinov (1820), nom. cons.
or some of these small clades cannot, however, be
justified. Not only have the limits of Cactaceae been
Sarraceniaceae Dumort. (1829), nom. cons.
stable over the years, but inclusion of Anacampseros
and relatives (Anacampserotaceae) and Portulacaceae
Styracaceae DC. & Spreng. (1821), nom. cons. s.s. in Cactaceae (compatible with the best phyloge-
Symplocaceae Desf. (1820), nom. cons.
netic hypotheses) would yield a clade characterized
solely by multicellular axillary hairs. Similarly, inclu-
sion of these two groups and Talinaceae in Cactaceae
Theaceae Mirb. ex Ker Gawl. (1816), nom. cons.
would yield a clade characterized by parallelocytic
It was clear in APG II that Theaceae s.l. could not
stomata (probably) and fruit characters, but the latter
be maintained. Subsequent work on the potential
have been subsequently lost in Cactaceae (Ogburn &
segregates has clarified the morphological pattern of
Edwards, 2009). Anacampserotaceae has only recently
variation (Stevens, 2001, for a summary). Sladeni-
been described (Nyffeler & Eggli, in press), recognition
aceae are recognized as distinct from Pentaphyla-
is compatible with their phylogenetic position.
caceae; although the two are sister taxa, they share
It has long been recognized that the limits
few obvious characters, and little would be gained by
of Molluginaceae s.l. are unclear (e.g. Endress &
uniting them. However, Ternstroemiaceae have much
Bittrich, 1993). Limeaceae and Lophiocarpaceae are
in common with Pentaphylacaceae and so the former
are included in the latter. Theaceae s.s. are not imme-
findings (Cuénoud et al., 2002). Molluginaceae s.l. are
yet another group much in need of basic anatomical,
The monogeneric Pellicieraceae are included in
developmental and phytochemical study. We still
Tetrameristaceae; the resulting family, with three
know little about relationships of Phytolaccaceae,
genera, is moderately well characterized. Mitrastemo-
almost certainly not a monophyletic family as cur-
naceae is a morphologically distinctive holoparasitic
rently circumscribed. However, its division, as in
family that is well embedded in Ericales.
Judd et al. (2007), is premature.
The biggest problem for APG III was the question
of how to treat Primulaceae and their immediate
relatives, a closely related group that in the past has
often been recognized as a separate order. Although
Cornaceae Bercht. & J.Presl (1825), nom. cons.
Primulaceae and relatives are clearly in Ericales,
taxon limits in this group have been problematic.
Maesaceae are a monogeneric family necessitated by
Grubbiaceae Endl. ex Meisn., (1841), nom. cons.
the break-up of Myrsinaceae, as are a monogeneric
Hydrangeaceae Dumort. (1829), nom. cons.
Samolaceae Raf. by the break-up of Primulaceae (or
Hydrostachyaceae Engl. (1894), nom. cons.
Theophrastaceae would have to be extended, so
becoming less recognizable; see Källersjö, Bergqvist &Anderberg, 2000; Ståhl & Anderberg, 2004, for infor-
mation). The limits of Myrsinaceae were extended,
Actinidiaceae Engl. & Gilg. (1824), nom. cons.
and those of Primulaceae correspondingly restricted.
Balsaminaceae A.Rich. (1824), nom. cons.
Given that the limits of the two best-known families
Clethraceae Klotzsch (1851), nom. cons.
in the group, Myrsinaceae and Primulaceae, have
Cyrillaceae Lindl. (1846), nom. cons.
been substantially changed, apomorphies are hard to
Diapensiaceae Lindl. (1836), nom. cons.
recognize (less so for Maesaceae and Primulaceae
s.s.), and the group as a whole has numerous syna-
pomorphies and is easy to recognize so we extend the
Fouquieriaceae DC. (1828), nom. cons.
limits of Primulaceae (see also Mabberley, 2008),
Lecythidaceae A.Rich. (1825), nom. cons.
although we know that this move will not be univer-
Marcgraviaceae Bercht. & J.Presl (1820), nom. cons.
*Mitrastemonaceae Makino (1911), nom. cons. $Pentaphylacaceae Engl. (1897), nom. cons. (includ-
Polemoniaceae Juss. (1789), nom. cons. §Primulaceae Batsch ex Borkh. (1797), nom. cons.
§*Boraginaceae Juss. (1789), nom. cons. (including
Källersjö, Myrsinaceae R.Br., nom. cons., Theo-
Icacinaceae Miers (1851), nom. cons.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
Metteniusaceae H.Karst. ex Schnizl. (1860–1870)
Orobanchaceae Vent. (1799), nom. cons.
Oncothecaceae Kobuski ex Airy Shaw (1965)
Paulowniaceae Nakai (1949)Pedaliaceae R.Br. (1810), nom. cons.
The limits of Boraginaceae are drawn broadly.
Phrymaceae Schauer (1847), nom. cons.
Not only are the phylogenetic relationships within
§Plantaginaceae Juss. (1789), nom. cons.
the family still unclear, but as we know more
about relationships within its component clades, they
become less easy to distinguish (e.g. Gottschling et al.,
Scrophulariaceae Juss. (1789), nom. cons.
2005 for Cordioideae A.Gray). Molecular data suggest
that Hoplestigmataceae are to be included in Boragi-
naceae s.l., being placed in or near Cordioideae (K.
Wurdack, pers. comm.; V. Savolainen and M. Powell,
Verbenaceae J.St.-Hil. (1805), nom. cons.
pers. comm.); Hoplestigma Pierre is similar in inflo-rescence, ovary, pollen, etc. to Boraginaceae. Relation-
Note that relationships among many families in
ships of Boraginaceae s.l. and Vahliaceae remain
Lamiales, and to a certain extent also their limits,
unclear, in the former case despite the sequencing of
are still unclear. Some of us would prefer a vastly
the whole plastid genome (Moore et al., in press).
expanded circumscription of Scrophulariaceae, far
Three families, Icacinaceae, Metteniusaceae and
beyond what it has ever included, whereas others
Oncothecaceae, are to be placed in this general area
are not so inclined. The limits of Plantaginaceae
of the tree. Furthermore, genera that used to be
have been further restricted since APG II by the
included in Icacinaceae s.l. are also to be found here,
recognition of the family of small herbs with rather
although they do not group with Icacinaceae s.s.
distinctive stem anatomy and floral morphology
(e.g. Linderniaceae), and Thomandersia has been
Cassinopsis Sond. and Emmotum Ham. (= Emmota-
removed from Acanthaceae as the monogeneric
ceae Tiegh.). All these taxa show similarities to Gar-
ryales, and circumscription of that order could easily
be expanded to include them if phylogenetic relation-
Solanales Juss. ex Bercht. & J.Presl (1820)
ships warranted it. Revised family limits depend on
Convolvulaceae Juss. (1789), nom. cons.
Montiniaceae Nakai (1943), nom. cons.
Eucommiaceae Engl. (1907), nom. cons.
$Garryaceae Lindl. (1834), nom. cons. (including
Sphenocleaceae T.Baskerv. (1839), nom. cons.
Although Aucubaceae and Garryaceae (both mono-
generic) appear distinct, there are several apomor-
Gentianales Juss. ex Bercht. & J.Presl (1820)
Aquifoliaceae Bercht. & J.Presl (1820), nom. cons.
Apocynaceae Juss. (1789), nom. cons.
Gelsemiaceae Struwe & V.A.Albert (1995)
Gentianaceae Juss. (1789), nom. cons.
Loganiaceae R.Br. ex Mart. (1827), nom. cons. Leptaulus Benth., previously unplaced, is assigned
to Cardiopteridaceae (Kårehed, 2001).
§Acanthaceae Juss. (1789), nom. cons. Bignoniaceae Juss. (1789), nom. cons.
Byblidaceae Domin (1922), nom. cons.
Asteraceae Bercht. & J.Presl (1820), nom. cons.
Gesneriaceae Rich. & Juss. (1816), nom. cons.
Calyceraceae R.Br. ex Rich. (1820), nom. cons.
Lamiaceae Martinov (1820), nom. cons.
$Campanulaceae Juss. (1789), nom. cons. (includ-
*Linderniaceae Borsch, K.Müll., & Eb.Fisch. (2005)
Lentibulariaceae Rich. (1808), nom. cons.
Goodeniaceae R.Br. (1810), nom. cons.
Martyniaceae Horan. (1847), nom. cons.
Menyanthaceae Dumort. (1829), nom. cons.
Oleaceae Hoffmanns. & Link (1809), nom. cons.
Pentaphragmataceae J.Agardh (1858), nom. cons.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
Dipsacales Juss. ex Bercht. & J.Presl (1820)
$Stylidiaceae R.Br. (1810), nom. cons. (including
§Caprifoliaceae Juss. (1789), nom. cons. [including
cons., Linnaeaceae Backlund, Morinaceae Raf.,
Expansion of Stylidiaceae to include the mono-
generic Donatiaceae is supported by morphologyand geography, and the expanded Campanulaceae
have strong support in molecular studies and are
adopted here as it is widely preferred (Judd et al.,
2007; Mabberley, 2008). The expanded family is well
within Campanulaceae s.l. are still unclear (Tank &
characterized, but half the clades it includes are
Donoghue, in press), and a future attempt to recog-
poorly characterized morphologically.
nize Lobeliaceae might either result in a clade poorly
supported morphologically or entail the recognition of
Griseliniaceae J.R.Forst. & G.Forst. ex A.Cunn.
§Escalloniaceae R.Br. ex Dumort. (1829), nom. cons.
(including Eremosynaceae Dandy, Polyosmaceae
Pennantiaceae J.Agardh (1858)Pittosporaceae R.Br. (1814), nom. cons.
This is a heterogeneous group of genera that forms
§Torricelliaceae Hu (1934) (including Aralidiaceae
a well-supported clade, but one of uncertain position
Philipson & B.C.Stone, Melanophyllaceae Takht.
and within which relationships are poorly supported
(Tank & Donoghue, in press). It is likely to be sisterto Asterales or, more probably, to all campanulids
Expansion of Torricelliaceae to include Aralidiaceae
apart from Asterales and Aquifoliales. Even if sister
and Melanophyllaceae is reasonable. All three are
to Asterales, inclusion in that order would make the
monogeneric and poorly known. Nevertheless, they
latter distinctly more heterogeneous; separate ordinal
form a strongly supported clade (e.g. Lundberg, 2001;
Plunkett, 2001; Kårehed, 2002, 2003), and that theywere kept separate before was a simple oversight.
The recognition of Myodocarpaceae results from our
Bruniaceae R.Br. ex DC. (1825), nom. cons.
improved understanding of relationships of members
§Columelliaceae D.Don (1828), nom. cons. (includ-
An order is needed for the two families above.
Winkworth, Lundberg & Donoghue (2008) found some
support for a position sister to Asterales and Tank &
Cynomoriaceae Endl. ex Lindl. (1833), nom. cons.
Donoghue (in press) found stronger support for a
position sister to the Paracryphiales–Dipsacales–
Petenaea Lundell (possibly Malvales)
Columelliaceae are broadly circumscribed because
Desfontainiaceae have much in common with them;both families are Andean and monogeneric.
†Paracryphiales Takht. ex Reveal (1992)
Albach DC, Martínez-Ortega MM, Fischer MA, Chase
§Paracryphiaceae Airy Shaw (1965) (including
MW. 2004. A new classification of the tribe Veroniceae –
problems and a possible solution. Taxon 53: 429–452. APG. 1998. An ordinal classification for the families of flow-
ering plants. Annals of the Missouri Botanical Garden 85:
Although these three families are at first sight
strikingly different, they have several characters in
APG II. 2003. An update of the Angiosperm Phylogeny Group
common and form a strongly supported clade (Tank &
classification for the orders and families of flowering plants:
Donoghue, in press); all are monogeneric and from
APG II. Botanical Journal of the Linnean Society 141:
the southwestern Pacific. Combination is in order (see
also Myrtales, Crossosomatales). Tank & Donoghue
Applequist WL, Wallace RS. 2001. Phylogeny of the portu-
(in press) found 100% bootstrap support for a position
lacaceous cohort based on ndhF sequence data. Systematic
of Paracryphiales as sister to Dipsacales. Botany 26: 406–419.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121 Backlund A, Bremer K. 1998. To be or not to be – principles Davis CC, Webb CO, Wurdack KJ, Jaramillo CA,
of classification and monotypic plant families. Taxon 47: Donoghue MJ. 2005. Explosive radiation of Malpighiales
supports a Mid-Cretaceous origin of modern tropical rain
Barkman TJ, McNeal JR, Lim S-H, Coat G, Croom HB,
forests. American Naturalist 165: E36–E65. Young ND, dePamphilis CW. 2007. Mitochondrial DNA Davis CC, Latvis M, Nickrent DL, Wurdack KJ, Baum
suggests at least 11 origins of parasitism in angiosperms
DA. 2007. Floral gigantism in Rafflesiaceae. Science 315:
and reveals genomic chimerism in parasitic plants. BMCEvolutionary Biology 7: 248. Der J, Nickrent D. 2008. A molecular phylogeny of Santal- Brockington SF, Alexandre R, Ramdial J, Moore MJ,
aceae (Santalales). Systematic Botany 33: 107–116. Crawley S, Dhingra A, Hilu K, Soltis PS Soltis DE. Doust AN, Stevens PF. 2005. A reinterpretation of the 2009. Phylogeny of the Caryophyllales and the evolution of
staminate flowers of Haptanthus. Systematic Botany 30:
the perianth. International Journal of Plant Sciences 170: Ducousso M, Béna G, Bourgeois C, Buyck B, Eyssartier Chase MW, Fay MF, Devey DS, Maurin O, Rønsted N, G, Vincelette M, Rabévohitra R, Randrihasipara L, Davies J, Pillon Y, Petersen G, Seberg O, Tamura MN, Dreyfus B, Prin Y. 2004. The last common ancestor of Asmussen CB, Hilu K, Borsch T, Davis JI, Stevenson
Sarcolaenaceae and Asian dipterocarp trees was ectomycor-
DW, Pires JC, Givnish TJ, Sytsma KJ, Graham SW,
rhizal before the India–Madagascar separation, about 88
McPherson MA, Rai HS. 2006. Multi-gene analyses of
million years ago. Molecular Ecology 13: 231–236.
monocot relationships: a summary. In: Columbus JT, Friar
Endress ME, Bittrich V. 1993. Molluginaceae. In: Kubitzki
EA, Hamilton CW, Porter JM, Prince LM, Simpson MG,
K, Rohwer JG, Bittrich V, eds. The families and genera of
eds. Monocots: comparative biology and evolution (vol. 1,vascular plants. II. Flowering plants: dicotyledons, magno-excluding Poales). Claremont, CA: Rancho Santa Ana
liid, hamamelid and caryophyllid families. Berlin: Springer,
Chase MW, Reveal JL. 2009. A phylogenetic classification of Endress PK, Doyle JA. 2009. Reconstructing the ancestral
the land plants to accompany APG III. Botanical Journal of
angiosperm flower and its initial specializations. Americanthe Linnean Society 161: 122–127. Journal of Botany 96: 22–66. Chase MW, Reveal JL, Fay MF. 2009. A subfamilial clas- Entwisle TJ, Weston PH. 2005. Majority rules, when sys-
sification for the expanded asparagalean families, Amaryl-
tematists disagree. Australian Journal of Systematic Botany
lidaceae, Asparagaceae and Xanthorrhoeaceae. Botanical18: 1–6. Journal of the Linnean Society 161: 132–136. Friedman WE. 2008. Hydatellaceae are water lilies with Chase MW, Zmarzty S, Lledó MD, Wurdack KJ, Swensen
gymnospermous tendencies. Nature 453: 94–97. SM, Fay MF. 2002. When in doubt, put it in Flacourti- Goremykin VV, Viola R, Hellwig FH. 2009. Removal of
aceae: a molecular phylogenetic analysis based on plastid
noisy characters from chloroplast genome-scale data sug-
rbcL DNA sequences. Kew Bulletin 57: 141–181.
gests revision of phylogenetic placement of Amborella and
Cheek M. 2006. The validation of two new family names in Ceratophyllum. Journal of Molecular Evolution 68: 197–204.
Malvales: Durionaceae and Brownlowiaceae. Kew BulletinGottschling M, Miller JS, Weigend M, Hilger HH. 2005.
Congruence of a phylogeny of Cordiaceae (Boraginales)
Chen J-M, Chen D, Robert GW, Wang Q-F, Guo Y-H.
inferred from ITS1 sequence data with morphology, ecology,
2004a. Evolution of apocarpy in Alismatidae using phylo-
and biogeography. Annals of the Missouri Botanical Garden
genetic evidence from chloroplast rbcL sequence data. 92: 425–437. Botanical Bulletin of Academia Sinica 45: 33–40. Hall JC, Sytsma KJ, Iltis HH. 2002. Phylogeny of Cappa- Chen J-M, Robert GW, Wang Q-F. 2004b. Evolution of
raceae and Brassicaceae based on chloroplast sequence
aquatic life forms in Alismatidae: phylogenetic estimation
data. American Journal of Botany 89: 1826–1842.
from chloroplast rbcL sequence data. Israel Journal of PlantHall JC, Iltis HH, Sytsma KJ. 2004. Molecular phylo- Sciences 52: 323–329.
genetics of core Brassicales, placement of orphan genera
Cronquist A. 1981. An integrated system of classification of Emblingia, Forchammeria, Tirania, and character evolu-
flowering plants. New York: Columbia University Press.
tion. Systematic Botany 29: 654–669. Cuénoud P, Savolainen V, Chatrou LW, Powell M, Hansen DR, Dastidar SG, Cau Z, Penaflor C, Kuehl JV, Grayer RJ, Chase MW. 2002. Molecular phylogenetics of Boore JL, Jansen RK. 2007. Phylogenetic and evolution-
Caryophyllales based on nuclear 18S rDNA and plastid
ary implications of complete chloroplast genome sequences
rbcL, atpB, and matK DNA sequences. American Journal of
of four early-diverging angiosperms: Buxus (Buxaceae),
Botany 89: 132–144. Chloranthus (Chloranthaceae), Dioscorea (Dioscoreaceae),
Davis CC, Chase MW. 2004. Elatinaceae are sister to
and Illicium (Schisandraceae). Molecular Phylogenetics and
Malpighiaceae; Peridiscaceae belong to Saxifragales. Ameri-Evolution 45: 547–563. can Journal of Botany 91: 262–273. Haston E, Richardson JE, Stevens PF, Chase MW, Davis CC, Wurdack KJ. 2004. Host-to-parasite gene Harris DJ. 2009. LAPG III: a linear sequence of the fami-
transfer in flowering plants: phylogenetic evidence from
lies in APG III. Botanical Journal of the Linnean Society
Malpighiales. Science 305: 676–678. 161: 128–131.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121 Heywood VH, Brummitt RK, Culham A, Seberg O. 2007.
systematics in four orders of rosids, including a broad
Flowering plant families of the World. Kew: Royal Botanic
survey of floral mucilage cells. Plant Systematics andEvolution 260: 199–221. Iles W, Smith SY, Graham SW. 2009. Robust resolution of Matthews ML, Endress PK. 2008. Comparative floral struc-
the backbone of Alismatales phylogeny. In Botany and
ture and systematics in Chrysobalanaceae s.l. (Chrysobal-
[http://2009.botanyconference.org/engine/search/
anaceae, Dichapetalaceae, Euphroniaceae, Trigoniaceae;
Malpighiales). Botanical Journal of the Linnean SocietyJansen RK, Cai Z, Raubeson LA, Daniell H, dePamphilis 157: 249–309. CW, Leebens-Mack J, Müller KF, Guisinger-Bellian M, Merckx V, Bidartondo MI. 2008. Breakdown and delayed Haberle RC, Hansen AK, Chumley TW, Lee S-B, Peery
cospeciation in the arbuscular mycorrhizal mutualism. Pro-R, McNeal JR, Kuehl JV, Boore JL. 2007. Analysis of 81 ceedings of the Royal Society, Series B 275: 1029–1035.
genes from 64 chloroplast genomes resolves relationships in
Merckx V, Schols V, Maas-van de Kamer H, Maas P, Huysmans S, Smets E. 2006. Phylogeny and evolution of
patterns. Proceedings of the National Academy of Sciences
Burmanniaceae (Dioscoreales) based on nuclear and mito-
of the United States of America 104: 19369–19374.
chondrial data. American Journal of Botany 93: 1684–1698. Jian S, Soltis PS, Gitzendanner MA, Moore MJ, Li R, Merckx V, Bakker FT, Huysmans S, Smets E. 2009. Bias Hendry TA, Qiu Y-L, Dhingra A, Bell C, Soltis DE.
and conflict in phylogenetic inference of myco-heterotrophic
2008. Resolving an ancient, rapid radiation in Saxifragales.
plants: a case study in Thismiaceae. Cladistics 25: 64–77. Systematic Biology 57: 38–57. Moore MJ, Bell CD, Soltis PS, Soltis DE. 2007. Using Judd WS, Campbell CS, Kellogg EA, Stevens PF, Dono-
plastid genome-scale data to resolve enigmatic relationships
ghue MJ. 2007. Plant systematics: a phylogenetic approach,
among basal angiosperms. Proceedings of the NationalAcademy of Sciences of the United States of America 104: Källersjö M, Bergqvist G, Anderberg AA. 2000. Generic
realignment in primuloid families of the Ericales s.l.: a
Moore MJ, Bell CD, Soltis PS, Soltis DE. 2008. Analysis
phylogenetic analysis based on DNA sequences from three
of an 83-gene, 86-taxon plastid genome data set resolves
chloroplast genes and morphology. American Journal of
relationships among several deep-level eudicot lineages. Botany 87: 1325–1341.
In: Botany 2008. [Botanical Society of America, http://
Kårehed J. 2001. Multiple origin of the tropical forest tree
2008.botanyconference.org / engine / search / index.php ? func=
family Icacinaceae. American Journal of Botany 88: 2259– Moore MJ, Soltis PS, Bell CD, Burleigh JG, Soltis DE. Kårehed J. 2002. Evolutionary studies in asterids emphasis- 2009. Phylogenetic analysis of 83 plastid genes resolves ing euasterids II. Uppsala: Acta Universitatis Upsaliensis.
relationships among major clades of eudicot angiosperms
Kårehed J. 2003. The family Pennantiaceae and its relation-
and reveals multiple rapid radiations. Proceedings of the
ship to Apiales. Botanical Journal of the Linnean SocietyNational Academy of Sciences of the United States of141: 1–24. Kubitzki K, Chase MW. 2002. Introduction to Malvales. In: Nickrent DL. 2002. Orígenes filogenéticos de las plantas
Kubitzki K, ed. The families and genera of vascular plants.
parásitas. In: López-Sáez JA, Catalán P, Sáez L, eds. V. Flowering plants. Dicotyledons. Malvales, CapparalesPlantas parásitas de la península Ibérica e islas Baleares. and non-betalain Caryophyllales. Berlin: Springer, 12–16. Lundberg J. 2001. Phylogenetic studies in the euasterids II Nickrent DL. 2007. Cytinaceae are sister to Muntingiaceae with particular reference to Asterales and Escalloniaceae.
(Malvales). Taxon 56: 1129–1135.
Uppsala: Acta Universitatis Upsaliensis. Nickrent DL, Der JP, Anderson FE. 2005. Discovery of the Mabberley DJ. 2008. Mabberley’s plant book: a portable
photosynthetic relatives of the ‘Maltese mushroom’ Cynomo-dictionary of plants, their classifications, and uses, 3rd edn. rium. BMC Evolutionary Biology 5: 38. Available at http://
Cambridge: Cambridge University Press. Malécot V, Nickrent DL. 2008. Molecular phylogenetic rela- Nyffeler R. 2007. The closest relatives of cacti: insights from
tionships of Olacaceae and related Santalales. Systematic
phylogenetic analyses of chloroplast and mitochondrial
Botany 33: 97–106.
sequences with special emphasis on relationships in the
Matthews ML, Endress PK. 2005a. Comparative floral
tribe Anacampseroteae. American Journal of Botany 94:
structure and systematics in Celastrales (Celastraceae,
Parnassiaceae, Lepidobotryaceae). Botanical Journal of theNyffeler R, Eggli U. In press. Disintegrating Portulacaceae Linnean Society 149: 129–194.
– a new familial classification of the suborder Portulacineae
Matthews ML, Endress PK. 2005b. Comparative floral
(Caryophyllales) based on molecular and morphological evi-
structure and systematics in Crossosomatales (Crossoso-
Ogburn RM, Edwards EJ. 2009. Anatomical variation in
Geissolomataceae, Ixerbaceae, Strasburgeriaceae). Botani-
Cactaceae and relatives: trait lability and evolutionary inno-
cal Journal of the Linnean Society 147: 1–46.
vation. American Journal of Botany 96: 391–408. Matthews ML, Endress PK. 2006. Floral structure and Oh S-H, Potter D. 2006. Description and phylogenetic posi-
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
tion of a new angiosperm family, Guamatelaceae, inferred
VI. Flowering plants. Dicotyledons. Celastrales, Oxalidales,
from chloroplast rbcL, atpB, and matK sequences. System-Rosales, Cornales, Ericales. Berlin: Springer, 255–257, 266–
atic Botany 31: 730–738. Pfeil BE, Crisp MD. 2005. What to do with Hibiscus? A Stevens PF. 2001 onwards. Angiosperm phylogeny website.
proposed nomenclatural resolution for a large and well-
Available at http://www.mobot.org/MOBOT/research/APweb/
known genus of Malvaceae and comments on paraphyly. Takhtajan A. 1997. Diversity and classification of flowering Australian Journal of Systematic Botany 18: 49–60. plants. New York: Columbia University Press. Plunkett GM. 2001. Relationship of the order Apiales to Tank DC, Donoghue MJ. In press. Phylogeny and phylo-
subclass Asteridae: a re-evaluation of morphological char-
genetic nomenclature of the Campanulidae based on an
acters based on insights from molecular data. Edinburgh
expanded sample of genes and taxa. Systematic Botany. In
Journal of Botany 8: 183–200. Refulio-Rodriguez NF, Olmstead R. 2008. Lamiales phy- Tamura MN, Yamashita J, Fuse S, Haraguchi M. 2004.
logeny. In: Botany 2008. [http://2008.botanyconference.org/
Molecular phylogeny of monocotyledons inferred from com-
engine/search/index.php?func=detail&aid=498]
bined analysis of plastid matK and rbcL gene sequences. Reveal J. 2008 onwards. A checklist of family and supra- Journal of Plant Research 117: 109–120.
familial names for extant vascular plants. Available at
Van Beusekom-Osinga RJ, van Beusekom CF. 1975.
http://www.plantsystematics.org/reveal/pbio/fam/
(Myrtales). Blumea 22: 255–266. Rudall PJ, Remizowa MV, Beer AS, Bradshaw E, Wang H, Moore MJ, Soltis PS, Bell CD, Brockington SF, Stevenson DW, Macfarlane TD, Tuckett RE, Yadav SR, Alexandre R, Davis CC, Latvis M, Manchester SR, Sokoloff DD. 2008. Comparative ovule and megagameto- Soltis DE. 2009. Rosid radiation and the rapid rise of
phyte development in Hydatellaceae and water lilies reveal
angiosperm-dominated forests. Proceedings of the National
a mosaic of features among the earliest angiosperms. Academy of Sciences of the United States of America 106: Annals of Botany 101: 941–956. Saarela JM, Rai HS, Doyle JA, Endress PK, Mathews S, Winkworth RC, Lundberg J, Donoghue MJ. 2008. Marchant AD, Briggs B, Graham SW. 2007. Hydatel-
Towards a resolution of campanulid phylogeny, with special
laceae identified as a new branch near the base of the
reference to the placement of Dipsacales. Taxon 57: 53–65.
angiosperm phylogenetic tree. Nature 446: 312–315. Worberg A, Alford MH, Quandt D, Borsch T. 2009. Simmons MP. 2004. Celastraceae. In: Kubitzki K, ed. The
Huerteales sister to Brassicales plus Malvales, and newly
families and genera of vascular plants. VI. Flowering plants.
circumscribed to include Dipentodon, Gerrardina, Huertea,Dicotyledons. Celastrales, Oxalidales, Rosales, Cornales,Perrottetia, and Tapiscia. Taxon 58: 468–478. Ericales. Berlin: Springer, 29–64. Wortley AH, Harris DJ, Scotland RW. 2007. On the Sosa V, Chase MW. 2003. Phylogenetics of Crossosomata-
taxonomy and phylogenetic position of Thomandersia.
ceae based on rbcL sequence data. Systematic Botany 28: Systematic Botany 32: 415–444. Wurdack KJ, Davis CC. 2009. Malpighiales phylogenetics: Soltis DE, Clayton JW, Davis CC, Gitzendanner MA,
gaining ground on one of the most recalcitrant clades in the
Cheek M, Savolainen V, Amorin AM, Soltis PS. 2007a.
angiosperm tree of life. American Journal of Botany 96:
Phylogenetic relationships of the enigmatic amphitropical
family Peridiscaceae. Taxon 56: 65–73. Zhang L-B, Simmons MP. 2006. Phylogeny and delimitation Soltis DE, Gitzendanner MA, Soltis PS. 2007b. A 567-
of the Celastrales inferred from nuclear and plastid genes.
taxon data set for angiosperms: The challenges posed by
Systematic Botany 31: 122–137.
Bayesian analyses of large data sets. International JournalZhang Z-H, Li C-Q, Li J. 2009. Phylogenetic placement of of Plant Sciences 168: 137–157. Cynomorium in Rosales inferred from sequences of the
Ståhl B, Anderberg AA. 2004. Maesaceae, Myrsinaceae. In:
inverted repeat region of the chloroplast genome. Journal of
Kubitzki K, ed. The families and genera of vascular plants.Systematics and Evolution 47: 297–304.
2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 105–121
KEPPNISREGLUR FYRIR SKÍÐAGÖNGU EFNISYFIRLIT 338 Þjálfun í brautum og lokun brauta 341 Starfsmenn og aðrir meðan á keppni stendur 372 Tæknilegur útbúnaður og undirbúningur 373 Mótaskrá-undirbúningur brauta-flokkaskipting 374 Skiptisvæðið 300 Skíðagöngukeppnir Skipulag 301 Mótsstjórn (organisesjons). 301.1. Fyrir stærri keppnir skal mótshaldari s
“ RELACION ENTRE EL TRATAMIENTO ANTIGIARDIÁSICO Y ESTADO NUTRICIONAL EN PREESCOLARES DEL ASENT AMIENTO HUMANO ALTO TRUJILLO DEL DISTRITO EL PORVENIR” Autores: Bazán Cayetano Johana, García Saucedo Sheyla, Campos Florián Julio Cátedra de Farmacología, Escuela de Farmacia y Bioquímica, Facultad de Farmacia y Bioquímica, Universidad Nacional de Trujillo, Perú. El present
 MagnolialesLauralesChloranthales†CommelinalesZingiberalesPoales
DasypogonaceaeAsparagalesLilialesPandanalesDioscorealesPetrosaviales†AlismatalesAcoralesCeratophyllales†RanunculalesSabiaceaeProteales
Buxales†Trochodendrales†GunneralesCucurbitalesFagalesRosalesFabalesCelastrales
Picramniales†CrossosomatalesMyrtalesGeranialesVitales†SaxifragalesDilleniaceaeBerberidopsidales†SantalalesCaryophyllalesCornalesEricales
SolanalesBoraginaceaeAquifolialesEscalloniales†AsteralesDipsacales
Figure 1. Interrelationships of the APG III orders and some families supported by jackknife/bootstrap percentages
MagnolialesLauralesChloranthales†CommelinalesZingiberalesPoales
DasypogonaceaeAsparagalesLilialesPandanalesDioscorealesPetrosaviales†AlismatalesAcoralesCeratophyllales†RanunculalesSabiaceaeProteales
Buxales†Trochodendrales†GunneralesCucurbitalesFagalesRosalesFabalesCelastrales
Picramniales†CrossosomatalesMyrtalesGeranialesVitales†SaxifragalesDilleniaceaeBerberidopsidales†SantalalesCaryophyllalesCornalesEricales
SolanalesBoraginaceaeAquifolialesEscalloniales†AsteralesDipsacales
Figure 1. Interrelationships of the APG III orders and some families supported by jackknife/bootstrap percentages