Plant Physiology and Biochemistry 43 (2005) 567–572
A CFTR chloride channel activator prevents HrpN -induced cell death
in Arabidopsis thaliana suspension cells
David Reboutier a,b,*, Cécile Frankart a, Régine Vedel b, Mathias Brault a, Ronald G. Duggleby c,
Jean Pierre Rona a, Marie Anne Barny b, François Bouteau a
a LEM, EA 3514, Université Paris 7, Case 7069, 2, place Jussieu, 75251 Paris cedex 5, France
b Laboratoire de Pathologie Végétale, UMR 217 Inra-INA-Paris VI, INA-PG, 16, rue Claude Bernard, 75231 Paris cedex 5, France
c Department of Biochemistry and Molecular Biology, The University of Queensland, Brisbane, Qld. 4072, Australia
Received 6 January 2005; accepted 17 March 2005
Abstract Erwinia amylovora is a necrogenic bacterium that causes fire blight of the Maloideae subfamily of Roseacae, such as apple and pear. It
provokes necrosis in aerial parts of susceptible host plants and the typical hypersensitive reaction in non-host plants. The secreted harpin,HrpN , is able by itself to induce an active cell death in non-host plants. Ion flux modulations were shown to be involved early in such
processes but very few data are available on the plasma membrane ion channel activities responsible for the pathogen-induced ion fluxes. Weshow here that HrpN
induces cell death in non-host Arabidopsis thaliana suspension cells. We further show that two cystic fibrosis trans-
membrane conductance regulator modulators, glibenclamide and bromotetramisole, can regulate anion channel activities and HrpN -induced
cell death. 2005 Elsevier SAS. All rights reserved. Keywords: Anion channel; Arabidopsis thaliana; Cell death; CFTR; Harpin
1. Introduction
membrane potential and ion flux variations are among theearliest signaling events detectable in response to pathogens
Erwinia amylovora is a bacterial pathogen that causes fire
and elicitors However, the underlying ion chan-
blight disease of apple, pear and other members of the
nel activities have been recorded rarely. Indeed, most of the
Rosaceae, its host plants. It secretes the HrpN
electrophysiological studies on plant cells are performed with
“hypersensitive response” (HR) elicitor HR cell death
patch-clamp technique applied to protoplasts and it seems
is a response of non-host plant to pathogen attack and con-
that the cell wall removing (protoplasts preparation) alters
sists of a rapid necrosis at the site of infection that cordons
the cell response capacity to pathogen or elicitor
off the pathogen and limits its spread There is a grow-
Another approach to analysis of ion channels in intact cells
ing consensus that HR is similar to animal programmed cell
that retains their cell wall is the microelectrode voltage-
death (PCD) and that ion channel regulation is a necessary
clamp technique. This technique allows long-term recording
of the free running membrane potential and whole cell ioncurrents, the internal medium remaining physiological (com-position non-controlled). We have shown previously, using
single microelectrode voltage-clamp (DSEVC), that Arabi-
AHAS, acetohydroxyacid synthase; AVD, apoptosis
volume decrease; CFTR, cystic fibrosis transmembrane conductance regu-
dopsis thaliana suspension cells respond to the fungal elici-
lator; DMSO, dimethyl sulfoxide; DSEVC, discontinuous single electrode
tor hypaphorine in the same way as root hairs, its natural tar-
voltage-clamp; HR, hypersensitive response; KORC, K+ outward rectifying
get Other studies demonstrated that suspension cultured
current; MAPK, mitogen activated protein kinase; PCD, programmed cell
cells are a powerful system of reduced complexity to analyze
the signal transduction pathway induced by pathogens
* Corresponding author. Tel.: +33 1 44 27 60 57; fax: +33 1 44 27 78 13. E-mail address: [email protected] (D. Reboutier).
Thus, we used A. thaliana suspension cells to inves-
0981-9428/$ - see front matter 2005 Elsevier SAS. All rights reserved. doi:10.1016/j.plaphy.2005.03.003
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
tigate early signaling events induced by HrpN . We showed
rents suggesting indirect effects on the channel proteins. Regulation of anion fluxes was reported in pathogen-inducedplant cell death. For example, a large nitrate efflux is neces-sary to induce cell death in tobacco in response to cryptogeinMoreover, anion channel antagonists have beenshown to interfere with elicitor or pathogen-induced responsessuch as Ca2+ influx production of active oxygen speciesMAPK activation and phytoalexin synthesisIn animal cells, Maeno et al. showed that apop-tosis volume decrease (AVD: cell shrinkage) is a major hall-mark of PCD. This AVD is due to a strong activation of ioneffluxes. In plant cells, the increase of anion effluxes inresponse to pathogen elicitors is consistent with theAVD. However, in A. thaliana suspension cells, we observeda decrease in anion current (efflux decrease) in response toHrpN
These data did not fit with the observations
described above but are closely related to those reported forhepatoblastoma apoptosis In this last model, apoptosisis induced by the decrease in cystic fibrosis transmembraneconductance regulator (CFTR) anion currents (members ofABC transporter superfamily). In plant, CFTR modulators,were shown to be effective on slow anion channels of Viciafaba guard cells Moreover, Leonhardt et al. showed,using antibodies, that slow anion channels are, or are closely,controlled by a polypeptide exhibiting an epitope shared with
Fig. 1. Effect of glibenclamide on anion currents.
the mammalian CFTR. Lastly, AtMRP5, a protein of the ABC
(A) Anion currents measured under control conditions and after adding 10 µMglibenclamide in the culture medium. Voltage pulses were –200 mV. Hol-
transporter superfamily, which has a high similarity to CFTR
ding potential was V . (B) Dose-dependent decrease of anion currents as a
and which is sensitive to glibenclamide, was suggested to con-
function of glibenclamide concentrations. Data represent the maximal effect.
trol ion channels In this study, our aim was to investi-
They were obtained from at least three independent experiments and were
gate the putative involvement of the anion current decrease in
fitted by a 5-parameter double exponential. The error bar corresponds to onestandard error.
HrpN -induced cell death on A. thaliana suspension cells
(non-host plant) by using CFTR modulators, glibenclamide,
cell CFTR-Cl– currents are inhibited at a half-maximal
a sulfonylurea (SU) molecule, as an inhibitor and bromotet-
concentration of 20 µM apoptosis is induced at 1 mM
ramisole as an activator, according to the hypothesis devel-
Thus, we tested the effect of different concentrations of
glibenclamide on anion currents. The deactivating currentspreviously characterized as anion currentsare sensitive to glibenclamide in a dose-dependent
2. Results
manner The glibenclamide concentration for half-maximum inhibition of anion current observed in our model
HrpN at 5 µg ml–1 (0.13 µM), a classically used HrpN
is about 7 µM of the same order of magnitude as
concentration increases cell death in comparison with
those observed for slow-type anion currents on plant cells
the cells treated with negative control According to
Thus, we tested the effect of 10 µM glibenclamide on
our hypothesis we thus tested in our model the effect of glib-
cell death. After 24 h, glibenclamide increased cell death,
enclamide and bromotetramisole. In animal systems, the con-
mimicking the HrpN -induced cell death Although
centrations of glibenclamide tested are highly variable: whole-
glibenclamide is effective on anion currents in our modeland in V. faba guard cells we checked if the
glibenclamide-induced cell death might be due to other known
Modulation of HrpN -induced cell death by anion channel modulators
glibenclamide or SU effects, i.e. K+ channel inhibition
Increase in cell death after a 24 h treatment with glibenclamide, HrpN alone
or herbicidal activity Glibenclamide (10 µM) does not
or mixed with bromotetramisole. Variations are given as a percentage withrespect to the control level. Data correspond to mean values ± S.D. and n is
induce a change in K+ outward rectifying current (KORC)
activity of A. thaliana suspension cells (D I
state for a + 80 mV voltage step = 1.3 ± 9%, n = 6, data not
shown). Yet in plants, the herbicidal activity of SU family
Cell death (%) 21 ± 3 (n = 6) 26 ± 4 (n = 3)
acts through AHAS inhibition AHAS catalyses the for-
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
Fig. 2. Effect of glibenclamide on AHAS activity. Inhibition of A. thaliana AHAS activity. Duplicate measurements were madeat each inhibitor concentration and the graph shows all values as separatepoints. From the data shown, the inhibition constant (Ki) was determined. The arrow indicates the AHAS inhibition at 10 µM.
mation of 2-acetolactate and 2-aceto-2-hydroxybutyrate asthe first step in the biosynthesis of the branched-chain aminoacids
glibenclamide-induced cell death involves this type of herbi-cidal effect or acts through ion channel regulation, we havetested whether glibenclamide could inhibit AHAS from A. thaliana. The residual AHAS activity at 10 µM glibencla-mide, compared to the corresponding control, was greater than95% for A. thaliana AHAS. The inhibition constant (Ki) cal-culated for AHAS, 146 ± 7 µM is much higher thanthose of known herbicidal sulfonylureas which have inhibi-
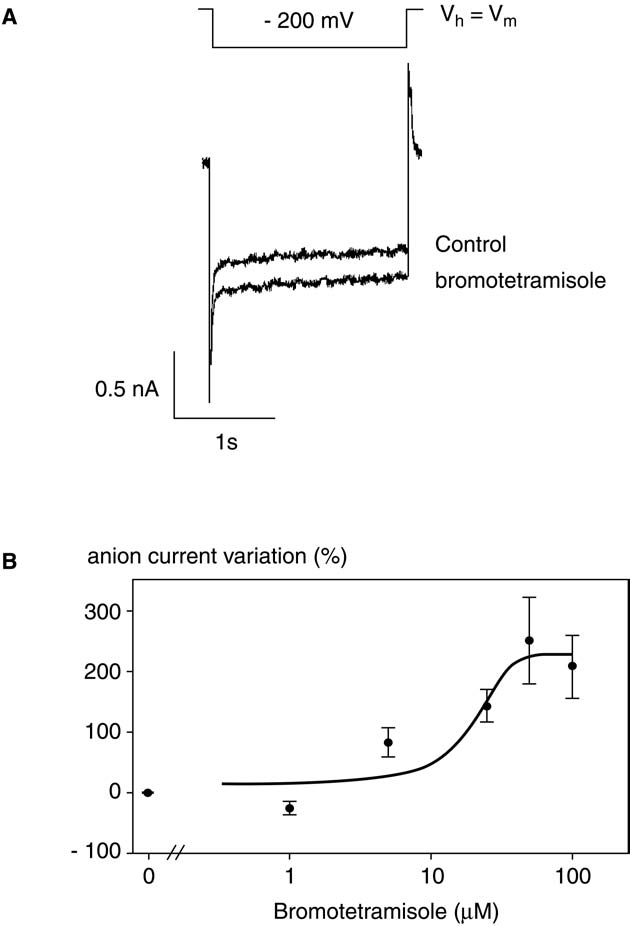
Fig. 3. Effect of bromotetramisole on anion currents.
tion constants in the 10–100 nM range for A. thaliana AHAS
(A) Anion currents measured under control conditions and after adding 5 µM
bromotetramisole in the culture medium. Voltage pulses were –200 mV. Hol-ding potential was V . (B) Dose-dependent increase of anion currents as a
In contrast to glibenclamide, the anion currents down regu-
function of bromotetramisole concentrations. Data represent the maximal
lated by HrpN are stimulated by bromotetramisole in a
effect. They were obtained from at least three experiments and were fitted by
dose-dependent manner The bromotetramisole half-
a 5-parameter double exponential. The error bar corresponds to one standard
maximum activation concentration we observed for anion cur-
rent in our model is about 20 µM Bromotetrami-
sole appeared thus as a useful tool to check if the increase of
ever, this decrease fits well with the PCD observed on hepa-
anion efflux could interfere with HrpN -induced cell death.
toblastoma cells by Kim et al. These authors showed
We tested bromotetramisole at 5 µM a low concentration that
that a decrease of CFTR anion current is a necessary step of
is able to increase anion currents. At this concentration, bro-
PCD development. To test the hypothesis that a decrease in
motetramisole counteracted the HrpN -induced cell death
anion current is involved in cell death, we used CFTR modu-
lators. Glibenclamide is believed to bind the CFTR channelallowing its block Glibenclamide is also a well-knowninhibitor of ATP-dependent K+ channels used as a therapeu-
3. Discussion
tic agent to treat type 2 diabetes If the increase in KORCinduced by HrpN was involved in cell death, the puta-
tive inhibition of KORC by glibenclamide should favor sur-
death symptom of aggression by an avirulent pathogen.
vival of cells. However, in our model, glibenclamide is effec-
We showed that HrpN could induce cell death in A. thaliana
tive on anion currents but failed to block the KORC.
suspension cells, a model non-host plant. The extent of
Glibenclamide also did not inhibit significantly AHAS
HrpN -induced cell death was in the same range as observed
and thus failed to induce any herbicidal effect as reported for
with another harpin, HrpZ, on A. thaliana suspension cells
other SU. Thus, the glibenclamide-induced cell death of A.thaliana suspension cells, mimicking the HrpN -induced cell
anion currents (anion effluxes) This result is not in accor-
death, most likely involves the inhibition of anion channels.
dance with previous studies suggesting that anion effluxes
The protecting effect of bromotetramisole (5 µM) against the
are a part of the pathogen or elicitor induced responses
lethal effect of HrpN reinforces this hypothesis. Bromotet-
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
ramisole is an uncompetitive inhibitor specific for alkaline
both preparations were compared on a 12% SDS-
phosphatase and tyrosine phosphatase. It inhibits a constitu-
polyacrylamide gel and quantified using the method of Brad-
tive, membrane associated, phosphatase activity and stabi-
represent 75% of the total protein prepara-
lizes the phosphorylated form of CFTR, thus increasing the
activity of CFTR chloride channel In our model, thestimulation of anion current by bromotetramisole is compat-
ible with such effects since phosphorylation is required forcomplete activation of slow anion channel A putative
Cell cultures (25 ml) were incubated for 15 min with 0.05%
phosphorylated form of anion channels could avoid the
Evans blue after 24 h of treatment with an effector then
HrpN -induced decrease in anion current, thus counteract-
washed with 200 ml of deionized water to remove the excess
ing cell death. In conclusion, our data suggest that the anion
and unbound dye. Dye bound to dead cells was solubilized in
channel involved in HrpN -induced cell death is, or is con-
50% methanol with 1% SDS for 60 min at 50 °C and quan-
trolled by, a protein exhibiting analogy with the CFTR and
tified at 595 nm The data represent an increase in cell
highlight the importance of ion channel regulation during the
death determined with regard to basal cell death control level.
The control levels correspond to treatment with negative con-trol for HrpN, and/or the solvent used with other effectors,water for bromotetramisole and methanol for glibenclamide. 4. Methods
The final concentration of methanol in all assays was 0.1%(v/v). 4.1. Cell culture and voltage-clamp experiments4.4. Acetohydroxyacid synthase (AHAS) assaysA. thaliana L. (ecotype Columbia) suspension cells were
cultured at 24 ± 2 °C, under continuous white light (40 µE m–
A. thaliana AHAS was purified as described previously
Assays were performed at 30 °C in a mixture containing
2 s–1) with rotation shaking, in a 1-l round bottom flask con-taining 350 ml Gamborg culture medium (main ions after
50 mM pyruvate, 1 mM thiamine diphosphate, 10 mM MgCl2
4 days of culture: 9 mM K+, 11 mM NO – The pH of
and 10 µM flavin adenine dinucleotide in 100 mM potassium
the culture medium was maintained at 5.8. Cells were sub-
phosphate buffer (pH 7.8). A colorimetric assay was
cultured weekly by a 10-fold dilution. The experiments were
employed. Glibenclamide was dissolved in the organic sol-
vent DMSO and added to the assay to a maximum concen-
For electrophysiological measurements, the cells were
tration of 400 µM. The final concentration of DMSO in all
impaled in the culture medium as previously described
The microelectrode resistance was 40–50 MX whenfilled with 600 mM KCl. Individual cells were voltage-clamped using an Axoclamp 2B amplifier (Axon Instru-
Acknowledgements
ments, Foster City, CA, USA) for discontinuous single elec-trode voltage-clamp experiments Voltage and current
We would like to thank the referees for helpful remarks,
were digitized with a personal computer fitted with a Digi-
D. Expert and H. El-Maarouf for critical reading of the manu-
data 1320A acquisition board (Axon Instruments). The elec-
script. Y.T. Lee supplied the sample of A. thaliana AHAS.
trometer was driven by pClamp software (pCLAMP8, Axon
This work was supported by funds from Inra and from the
Instruments). Experiments were performed at 22 ± 2 °C.
MENRT to EA 3514 and ACI 5078. D. Reboutier was sup-ported by a grant from MENRT.
The harpin was prepared and partially purified as described
References
previously Briefly, Escherichia coli strain K38(pGp1-2; pMAB64) containing the cloned hrpN
M. Bradford, A rapid and sensitive method for quantification of
microgram quantities of protein utilizing the principle of dye-binding,
was grown up to an optical density of 0.4 at 600 nm. T7 RNA
polymerase was induced according to Tabor et al. The
F. Becq, B. Verier, X.B. Chang, J.R. Riordan, J.W. Hanrahan, cAMP-
cells were harvested, resuspended in 12 ml of phosphate buffer
and Ca2+-independent activation of cystic fibrosis transmembrane
(pH 7, 10 mM), boiled for 10 min and centrifuged. The super-
conductance regulator channels by phenylimidazothiazole drugs, J.
natant was recovered and concentrated using Centricon10
Biol. Chem. 271 (1996) 16171–16179.
(Amicon). To assess the contribution of E. coli protein back-
M. Brault, Z. Amiar, A.M. Pennarun, M. Monestiez, Z. Zhang, D. Cor-
ground to membrane response and cell death, a cell free prepa-
nel, O. Dellis, H. Knight, F. Bouteau, J.P. Rona, Plasma membranedepolarization induced by abscisic acid in Arabidopsis suspension
ration of E. coli strain K38 (pGp1-2; pT7-7), containing the
cells involves reduction of proton pumping in addition to anion chan-
pT7-7 vector without the hrpN insert, was prepared in paral-
nel activation, which are both Ca2+ dependent, Plant Physiol. 135
lel to harpin and used as negative control. Protein contents of
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
D.E. Carden, H.H. Felle, The mode of action of cell wall-degrading
of stomatal movements and guard cell potassium channels by sulfo-
enzymes and their interference with Nod factor signalling in Medi-
nylureas and potassium channel openers, Proc. Natl. Acad. Sci. USA
cago sativa root hairs, Planta 216 (2003) 993–1002.
A.K. Chang, R.G. Duggleby, Expression, purification and character-
[23] N. Leonhardt, A. Vavasseur, C. Forestier, ATP binding cassette modu-
ization of Arabidopsis thaliana acetohydroxyacid synthase, Biochem.
lators control abscisic acid-regulated slow anion channels in guard
cells, Plant Cell 11 (1999) 1141–1152.
A.K. Chang, R.G. Duggleby, Herbicide-resistant forms of Arabidop-sis thaliana acetohydroxyacid synthase: characterization of the cata-
[24] N. Leonhardt, I. Bazin, P. Richaud, E. Marin, A. Vavasseur, C. Fores-
lytic properties and sensitivity to inhibitors of four defined mutants,
tier, Antibodies to the CFTR modulate the turgor pressure of guard
cell protoplasts via slow anion channels, FEBS Lett. 494 (2001)15–18.
U. Conrath, C.M.J. Pieterse, B. Mauch-Mani, Priming in plant–patho-gen interactions, Trends Plant Sci. 5 (2002) 210–216.
[25] A. Levine, R.I. Pennell, M.E. Alvarez, R. Palmer, C. Lamb, Calcium-
J.L. Dangl, R.A. Dietrich, M.H. Richberg, Death don’t have no mercy:
mediated apoptosis in a plant hypersensitive disease resistance
cell death programs in plant–microbe interactions, Plant Cell 8 (1996)
response, Curr. Biol. 6 (1996) 427–437.
[26] W. Ligterink, T. Kroj, U. zur Nieden, H. Hirt, D. Scheel, Receptor-
J. Ebel, A.A. Bhagwat, E.G. Cosio, M. Feger, U. Kissel, A. Mithöfer,
mediated activation of a MAP kinase in pathogen defense of plants,
T. Waldmüller, Elicitor-binding proteins and signal transduction in the
activation of a phytoalexin defense response, Can. J. Bot. 73 (1995)506–510.
[27] E. Maeno, Y. Ishizaki, T. Kanaseki, A. Hazama, Y. Okada, Normotonic
cell shrinkage because of disordered volume regulation is an early
[10] H. El-Maarouf, M.A. Barny, J.P. Rona, F. Bouteau, Harpin, a hyper-
prerequisite to apoptosis, Proc. Natl. Acad. Sci. USA 97 (2000)
sensitive response elicitor from Erwinia amylovora, regulates ion
channel activities in Arabidopsis thaliana suspension cells, FEBSLett. 497 (2001) 82–84.
[28] R. Mannhold, KATP channel openers: structure–activity relationships
[11] A.S. Finkel, S. Redman, Theory and operation of a single microelec-
and therapeutic potential, Med. Res. Rev. 24 (2004) 213–266.
trode voltage clamp, J. Neurosci. Methods 11 (1984) 101–127.
[29] B. Naton, K. Hahlbrock, E. Schmelzer, Correlation of rapid cell death
[12] N. Gaedeke, M. Klein, U. Kolukisaoglou, C. Forestier, A. Müller,
with metabolic changes in fungus-infected, cultured parsley cells,
M. Ansorge, D. Becker, Y. Mammun, K. Kuchler, B. Schulz, B. Muel-
Plant Physiol. 112 (1996) 433–444.
ler-Roeber, E. Martinoia, The Arabidopsis thaliana ABC transporterAtMRP5 controls root development and stomata movement, EMBO J.
[30] T. Nürnberger, D. Nennstiel, T. Jabs, W.R. Sacks, K. Halbrock,
D. Scheel, High affinity binding of a fungal oligopeptide elicitor to
[13] O.L. Gamborg, R.A. Miller, K. Ojima, Nutrient requirements of
parsley plasma membranes triggers multiple defense responses, Cell
suspension cultures of soybean root cells, Exp. Cell Res. 50 (1968)
[31] Y. Okada, E. Maeno, Apoptosis, cell volume regulation and volume-
[14] S. Gaudriault, L. Malandrin, J.P. Paulin, M.A. Barny, DspA, an essen-
regulatory chloride channels, Comp. Biochem. Physiol. A Mol. Integr.
tial pathogenicity factor of Erwinia amylovora showing homology
with AvrE of Pseudomonas syringae, is secreted via the Hrp secretionpathway in a DspB-dependent way, Mol. Microbiol. 26 (1997) 1057–
[32] S.M. Pike, A.L. Adam, X.A. Pu, M.E. Hoyos, R. Laby, S.V. Beer,
A. Novacky, Effect of Erwinia amylovora harpin on tobacco leaf cellmembranes are related to leaf necrosis and electrolyte leakage and
[15] J.T. Greenberg, Programmed cell death: a way of life for plants, Proc.
distinct from perturbation caused by inoculated E. amylovora,
Natl. Acad. Sci. USA 93 (1996) 12094–12097.
Physiol. Mol. Plant Pathol. 53 (1998) 39–60.
[16] F.M. Hughes, C.D. Bortner, G.D. Purdy, J.A. Cidlowski, Intracellular
K+ suppresses the activation of apoptosis in lymphocytes, J. Biol.
[33] L.P. Popham, S.M. Pike, A. Novacky, The effect of harpin from
Erwinia amylovora on the plasmalemma of suspension-cultured
[17] T. Jabs, M. Tschöpe, C. Colling, K. Hahlbrock, D. Scheel, Elicitor-
tobacco cells, Physiol. Mol. Plant Pathol. 47 (1995) 39–50.
stimulated ion fluxes and O – from the oxidative burst are essential
[34] A. Pugin, J.M. Frachisse, E. Tavernier, R. Bligny, E. Gout, R. Douce,
components in triggering defense gene activation and phytoalexin
J. Guern, Early events induced by the elicitor cryptogein in tobacco
synthesis in parsley, Proc. Natl. Acad. Sci. USA 94 (1997) 4800–
cells: involvement of a plasma membrane NADPH oxidase and acti-
vation of glycolysis and the pentose phosphate pathway, Plant Cell 9
[18] J.A. Kim, Y.S. Kang, S.H. Lee, E.H. Lee, B.H. Yoo, Y.S. Lee, Glib-
enclamide induced apoptosis through inhibition of CFTR-Cl– chan-nels and intracellular Ca2+ release in HepG2 human hepatoblastoma
[35] V.K. Rajasekhar, C. Lamb, R.A. Dixon, Early events in the signal
cells, Biochem. Biophys. Res. Commun. 261 (1999) 682–686.
pathway for the oxidative burst in soybean cells exposed to avirulentPseudomonas syringae pv glycinea, Plant Physiol. 120 (1999) 1137–
[19] B. Klüsener, E.W. Weiler, Pore-forming properties of elicitors of plant
defense reactions and cellulolytic enzymes, FEBS Lett. 459 (1999)263–266.
[36] D. Reboutier, M. Bianchi, M. Brault, C. Roux, A. Dauphin, J.P. Rona,
[20] M. Krause, J. Durner, Harpin inactivates mitochondria in Arabidopsis
V. Legue, F. Lapeyrie, F. Bouteau, The indolic compound hypaphorine
suspension cells, Mol. Plant Microb. Interact. 17 (2004) 131–139.
produced by ectomycorrhizal fungus interferes with auxin action andevokes early responses in nonhost Arabidopsis thaliana, Mol. Plant
[21] E. Lam, Controlled cell death, plant survival and development, Nat.
Microbe Interact. 15 (2002) 932–938.
Rev. Mol. Cell. Biol. 5 (2004) 305–315.
[22] N. Leonhardt, E. Marin, A. Vavasseur, C. Forestier, Evidence for the
[37] M. Sasabe, K. Takeuchi, S. Kamoun, Y. Ichinose, F. Govers,
existence of a sulfonylurea-receptor-like protein in plants: modulation
K. Toyoda, T. Shiraishi, T. Yamada, Independent pathways leading to
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
apoptotic cell death, oxidative burst and defense gene expression in
[42] Z.M. Wei, R.J. Laby, C.H. Zumoff, D.W. Bauer, S.Y. He, A. Collmer,
response to elicitin in tobacco cell suspension culture, Eur. J. Bio-
S.V. Beer, Harpin, elicitor of the hypersensitive response produced by
the plant pathogen Erwinia amylovora, Science 257 (1992) 85–88.
[38] C. Schmidt, I. Schelle, Y.J. Liao, J.I. Schröeder, Strong regulation of
[43] D. Wendehenne, O. Lamotte, J.M. Frachisse, H. Barbier-Brygoo,
slow anion channels and abscisic acid signaling in guard cells by
A. Pugin, Nitrate efflux is an essential component of the cryptogein
phosphorylation and dephosphorylation events, Proc. Natl. Acad. Sci.
signaling pathway leading to defense responses and hypersensitive
cell death in tobacco, Plant Cell 14 (2002) 1937–1951.
[39] D.N. Sheppard, M.J. Welsh, Effect of ATP-sensitive K+ channel regu-
[44] B. Xu, B.A. Wilson, L. Lu, Induction of human myeloblastic
lators on cystic fibrosis transmembrane conductance regulator chlo-
ML-1 cell G1 arrest by suppression of K+ channel activity, Am. J.
ride currents, J. Gen. Physiol. 100 (1992) 573–591.
[40] B.K. Singh, M.A. Stidham, D.L. Shaner, Assay of acetohydroxyacid
synthase, Anal. Biochem. 171 (1988) 173–179.
[45] S. Zimmermann, J.M. Frachisse, S. Thomine, H. Barbier-Brygoo,
J. Guern, Elicitor-induced chloride efflux and anion channels in
[41] S. Tabor, in: F.A. Ausubel, R. Brent, R.E. Kingston, D.D. Moore,
tobacco cell suspension, Plant Physiol. Biochem. 36 (1998) 665–674.
J.A. Seidman, K. Struhl (Eds.), Current Protocols in Molecular Biol-ogy, vol. 3, Greene Publishing and Wiley Interscience, New York,
[46] Z. Zhou, S. Hu, T.C. Hwang, Probing an open CFTR pore with organic
anion blockers, J. Gen. Physiol. 120 (2002) 647–662.
MEDCO BY MAIL ORDER FORM Member Information Please verify or provide member information below. ❏ Please send me e-mail notices about the status of the Member ID: enclosed prescription(s) and online ordering at: Group: (Medco will keep this address on file for all orders fromthis membership until another shipping address isprovided by any person in this membership.
CAPP-INFO Bulletin d’information du CAPP N° 15, Novembre 2000 (Contact Avis Pharmacologique et Pharmaceutique) Bip : 8 65 60 MEDICAMENTS ET ALIMENTATION Avant ? Pendant ? Après le repas ? I. INTRODUCTION Les aliments peuvent influer sur l’efficacité et la tolérance des médicaments de différentes manières. La prise simultanée de nourriture peut ralentir

 Plant Physiology and Biochemistry 43 (2005) 567–572
A CFTR chloride channel activator prevents HrpN -induced cell death
in Arabidopsis thaliana suspension cells
David Reboutier a,b,*, Cécile Frankart a, Régine Vedel b, Mathias Brault a, Ronald G. Duggleby c,
Jean Pierre Rona a, Marie Anne Barny b, François Bouteau a
a LEM, EA 3514, Université Paris 7, Case 7069, 2, place Jussieu, 75251 Paris cedex 5, France
b Laboratoire de Pathologie Végétale, UMR 217 Inra-INA-Paris VI, INA-PG, 16, rue Claude Bernard, 75231 Paris cedex 5, France
c Department of Biochemistry and Molecular Biology, The University of Queensland, Brisbane, Qld. 4072, Australia
Received 6 January 2005; accepted 17 March 2005
Abstract
Plant Physiology and Biochemistry 43 (2005) 567–572
A CFTR chloride channel activator prevents HrpN -induced cell death
in Arabidopsis thaliana suspension cells
David Reboutier a,b,*, Cécile Frankart a, Régine Vedel b, Mathias Brault a, Ronald G. Duggleby c,
Jean Pierre Rona a, Marie Anne Barny b, François Bouteau a
a LEM, EA 3514, Université Paris 7, Case 7069, 2, place Jussieu, 75251 Paris cedex 5, France
b Laboratoire de Pathologie Végétale, UMR 217 Inra-INA-Paris VI, INA-PG, 16, rue Claude Bernard, 75231 Paris cedex 5, France
c Department of Biochemistry and Molecular Biology, The University of Queensland, Brisbane, Qld. 4072, Australia
Received 6 January 2005; accepted 17 March 2005
Abstract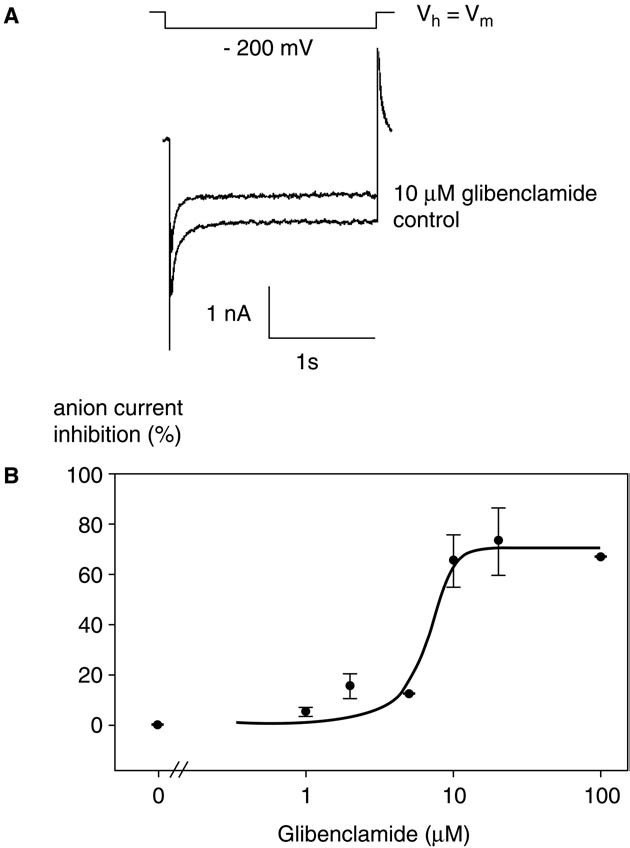 D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
tigate early signaling events induced by HrpN . We showed
rents suggesting indirect effects on the channel proteins.
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
tigate early signaling events induced by HrpN . We showed
rents suggesting indirect effects on the channel proteins.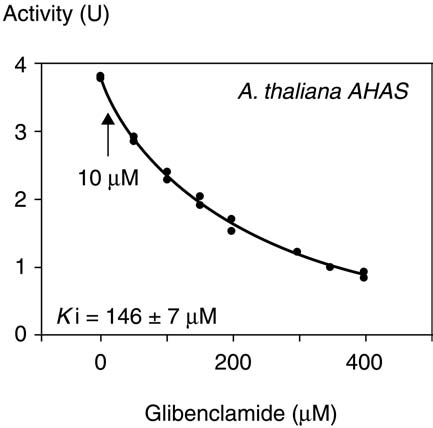
 D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
Fig. 2. Effect of glibenclamide on AHAS activity.
D. Reboutier et al. / Plant Physiology and Biochemistry 43 (2005) 567–572
Fig. 2. Effect of glibenclamide on AHAS activity.