Veterinary Parasitology 115 (2003) 185–198
Department of Molecular Microbiology, Washington University School of Medicine,660 S Euclid Box 8230, St. Louis, MO 63110, USAAbstract Toxoplasma gondii is a wide spread protozoan parasite belonging to the phylum Apicomplexa.
Members of this group of parasites cause economically and medically important diseases in a varietyof animals, including humans. T. gondii is notable among this group for the fact that it readily infectsvirtually all warm-blooded vertebrates, while most apicomplexans are typically restricted in theirhost range. Individual strains of T. gondii are genetically quite similar and molecular studies indicatethat the vast majority (>95%) of strains can be grouped into one of three distinct clonal lineages. Remarkably, these three lineages are comprised of different combinations of two parental alleles,indicating they arose from a single genetic cross. The unusual population structure of T. gondiioriginated within the last 10,000 years, while the genus itself is approximately 10 million years old. The remarkable success of the three lineages following their recent origin is likely attributable to adramatic change in the life cycle that greatly facilitated transmission. Prior to the cross that gave riseto the clonal lineages, and in all closely related parasites, transmission occurred by an obligatorytwo-host life cycle that alternated between definitive (carnivorous) and intermediate (herbivorous)hosts. The reassortment of genes that occurred in the cross allowed direct oral transmission betweenmany different intermediate hosts. These findings illustrate that complex biological life cycles canchange rapidly and dramatically, thus presenting a constant source of new zoonotic infections. 2003 Elsevier B.V. All rights reserved. Keywords: Virulence; Mutation; Life cycle; Protozoan parasite; Recombination; Evolution
1. Introduction to the Apicomplexa Toxoplasma gondii belongs to the phylum Apicomplexa, an ancient group of protozoans
that consists of an estimated 5000 species (The Apicomplexa are membersof a monophyletic group known as Alveolates, which also contains the ciliates and di-noflagelates This
∗ Tel.: +1-314-362-8873; fax: +1-314-362-3212. E-mail address: [email protected] (L. David Sibley).
0304-4017/03/$ – see front matter 2003 Elsevier B.V. All rights reserved. doi:10.1016/S0304-4017(03)00206-1
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
relationship is supported both by morphological comparisons (presence of sub-cortical alve-olar membranes) and by molecular analyses based on conserved protein sequences or smallsubunit (SSU) rRNA sequences. The phylum Apicomplexa contains organisms that shareapical specializations related to their polarized attachment and entry into host cells All extant lineages of the Apicomplexa are obligate intracellular para-sites. Despite lacking a fossil record, molecular phylogenies indicate that the origin of thisgroup spans some 1 billion years of time (Not surprisinglygiven this long history, the Apicomplexa currently inhabit a wide range of hosts includ-ing annelids, mollusks, insects, amphibians, reptiles, birds, and mammals Among the Apicomplexa there are several major clades represented by the piroplasms (e.g. Theileria, Babesia), haemosporinids (e.g. Plasmodium), gregarines and somewhat closelyrelated cryptosporidians (the enteric coccidia (e.g. Cyclospora, Eime-ria), and the tissue-cyst forming coccidia It is among this group that T. gondiiresides along with the animal pathogens Sarcocystis spp., Neospora spp., and the largelynon-pathogenic genera Hammondia and Besnoitia (The relative age since acommon ancestry of the tissue-cyst forming coccidia is approximately ∼250 million yearsago (mya) based on analyses of the SSU regions of the rRNA genes (Thistime period is similar to the expansion of the relevant mammalian hosts that are parasitizedby this group (suggesting that divergence of mammalian hosts was thedriving force for development of these distinct parasite lineages.
Coccidian life cycles come in two basic types. Monoxenous life cycles occur in a single
host species where they undergo asexual expansion and then sexual recombination withinthe gut, leading to shedding of a resistant spore-like stage (oocyst) into the environment. Transmission is exclusively fecal–oral and host range is generally quite restricted. In con-trast, heteroxenous life cycles alternate between sexual replication in a definitive host andasexual replication in a separate intermediate host. Among the tissue-cyst forming coccid-ians, heteroxenous life cycles typically involve a carnivorous definitive host that becomesinfected by ingesting tissues of an herbivorous intermediate host. Intermediate hosts becomeinfected by oocysts shed into the environment by the definitive host. The specific factorsthat control these different life cycle features are generally unknown; however, tremendousvariation exists in the range of intermediate hosts exploited by a given parasite. For example,while virtually all warm-blooded vertebrates can serve as intermediate hosts for T. gondiithe closely related parasite Neospora caninum exploits onlyungulates Speciation has resulted from an even finer divisionof intermediate hosts within the genus Sarcocystis where individual species typically infecta single genus of mammals (These examples highlight the enormous diver-gence of complex life cycle traits that occurs within this closely related group of parasites. 1.3. Rapid evolutionary changes in complex life cycles
The morphological similarities that were utilized in creating the Apicomplexa have
largely been borne out by molecular analyses (However, there are notable
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
exceptions, particularly in dealing with complex traits like life cycles or modes of trans-mission. For example, SSU phylogeny of the coccidian genus Carypspora places it close toEimeria and Cyclospora (both of which are exclusively monoxenous ina variety of host species. While most species of Carypospora are monoxenous, others arefacultatively heteroxenous (Likewise, while some members of the Isosporagenus are closely aligned phylogenetically with heteroxenous genera such as Toxoplasma,all known members of Isospora are monoxenous (The widespread occurrenceof heteroxenous life cycles within the tissue-cyst forming coccidians is suggestive that thislife style arose multiple independent times within this group during their history (Such a major life cycle change is likely to allow for rapid expansion within new hosts,and hence acquisition of this trait has a strong selective advantage. As a consequence, com-plex features such as life cycles may not be reliable indicators of historical origin. This con-cept has also been emphasized in studies of malarial parasites where the periodicity of fever,a trait influencing transmission, does not correlate well with molecular phylogeny (More broadly, phylogenetic analysis of cytochrome b gene sequences in dif-ferent malarial species reveals that classifications traditionally based on similarities in lifehistory traits do not accurately reflect common ancestry (
2. Population genetic structure of T. gondii
Early studies on T. gondii strains indicated that different isolates were remarkably similar
(The application of molecular approaches to strain typingrevealed that while strains of T. gondii are indeed highly similar, they fall into discreteclonal groups. Clonality is not an unusual feature among microorganisms and it has beenreported for many protozoan parasites previously (However, thelimited number of lineages and extensive linkage disequilibrium in T. gondii makes thisone of the most extreme examples of clonality. An even more perplexing finding is that thethree major strain types possess only two separate alleles at any given locus. Moreover, thealleles themselves are highly similar, being ∼98% identical at the nucleotide level. Thistruly remarkable population genetic pattern provides important clues about its origin, thegenetic forces driving success of the strains, and their underlying biological traits.
Initial studies of multi-locus enzyme electrophoresis revealed remarkably few alleles
exist at different loci in strains of T. gondii (Restriction fragmentlength polymorphism (RFLP) analysis of a larger set of isolates clearly distinguished asingle clonal group of strains that was acutely virulent in mice (Type I strains share a combination of alleles at many different loci, and this patternis different than that found in non-virulent strains. Virulent strains are defined as havingan LD100 of a single viable organism, while non-virulent strains typically have LD50s of
≥103 and readily establish chronic infection in mice (Further geneticprofiling by RFLP analyses indicated that the remaining non-virulent strains comprise twodiscrete clonal types known as types II and III (Type II strains arethe most common both in chronically infected animals and in human toxoplasmosis cases,while type III strains are largely confined to animals (
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
While it was widely appreciated that strains of T. gondii differed in virulence, this trait
was initially assumed to be due to passage history or environmental factors. This perspec-tive may have been influenced by early isolation of the virulent type I RH strain (which was then extensively propagated in the laboratory and widely used by manygroups. While a long passage history might lead to alterations in virulence, it is now clearthat acute virulence is primarily a consequence of genotype rather than propagation. Thisconclusion is supported by the fact that other type I strains (a distinction made based ongenotype) share the acute virulence phenotype even when analyzed as primary or low pas-sage isolates (Furthermore, the virulence of the differentstrain types in the mouse model is relatively stable over time (While repeated passage of strains has been reported to increase the virulenceof types II and III strains, they do not achieve the high lethality characteristic of type Istrains.
Despite the overwhelming clonality of T. gondii, some isolates show examples of mixed
genotypes, suggesting they are the products of naturally occurring recombination Furthermore, experimental crosses produced in the laboratory indicate thatstrains readily mate and undergo meiosis following classical Mendelian rules (). Why then is clonality maintainedas such a striking feature of natural populations? Several reasons have been given for thispattern including: (1) intermediate hosts are typically infected with only a single strainsuch that when ingested by a cat, the resulting infection is monotypic and does not leadto production of new genotypes, (2) direct transmission between successive intermediatehosts renders the cat non-essential in the life cycle, hence bypassing the sexual cycle (seebelow). This stands in marked contrast to malaria, where the definitive host (mosquito) playsan essential role in transmission. Hence, while clonality has been occasionally observedin malaria, it is confined to specific geographic outbreaks with low transmission rates andis not a general feature
3. Virulence and pathogenesis
The mouse model provides a convenient system to evaluate virulence and pathogenesis
during acute and chronic toxoplasmosis. Intraperitoneal inoculation provides the most re-liable, quantitative challenge, despite the fact that this does not represent a natural routeof infection. The more natural, oral challenge model has also been extensively explored toevaluate the contribution of host genetic factors to resistance Thesestudies have indicated an important role of MHC class II alleles as well as several other loci inmediating resistance (Certain genetic backgrounds, notably C57BL/6mice, develop high levels of intestinal necrosis following oral challenge and CNS disease following ip challenge (with thetype II strain ME49. A rat model has also been developed to assess the contribution of hostgenetic factors in resistance to infection
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
One of the most striking features of T. gondii is that the high virulence of type I strains
is independent of the genetic background of the mouse host (Challengeby type I strains always leads to lethal outcome and the minimal infectious dose is the sameas the minimal lethal dose, a situation that is quite unusual. In contrast, types II and IIIhave variable LD50s that depend on route of inoculation, host genotype and dose. Studiesin the mouse model indicate that the acute virulence of type I strains is due to a rapid andoverwhelming growth, leading to high tissue burdens of the parasite and the triggering of amassive TH1 type cytokine response (The extremely high levels of TH1 cytokines including IL-12, IL-18, IFN-␥, and TNF-␣likely contribute to tissue pathology and the ultimate demise of the animal. Paradoxically,these same cytokines are needed to control infections by non-virulent strains; however, inthis circumstance they are produced at appropriate levels and are down regulated followinginitial induction (
The combination of clonality evident in T. gondii with the lethality of type I strains
results in the appearance that many genes are linked with acute virulence. This should notbe construed as evidence that such genes participate in virulence directly however, as theseassociations may be purely fortuitous due to the high linkage disequilibrium. Fortunately,T. gondii is equipped with good techniques for both reverse and forward genetics (making it possible to directly test the role of putative virulence genes and topotentially fulfill Koch’s molecular postulates (
The large differences in acute virulence of T. gondii strains lends itself to quantitative trait
analysis, a genetic mapping technique that was developed to assess multigenic traits thathave a continuous rather than discrete distribution (This approach has recently been applied to examining the genetic basis ofacute virulence in T. gondii. A genetic cross between the acutely virulent type I strain GT-1and the non-virulent type III strain CTG was used to generate F1 recombinant progeny. These progeny were then tested to establish their virulence in the mouse model followingip challenge with tachyzoites. Separately, they were genotyped using a combination ofapproximately 70 polymorphic DNA markers that were analyzed by PCR amplificationfollowed by RFLP analysis. Analysis of the segregation of acute virulence revealed thatthis trait is multigenic and the key determinant is a locus on chromosome VII (It is estimated that this single locus may contribute as much as 50% of the trait. Additional loci, including one on chromosome IV are also implicated in acute virulence(Importantly, the alleles at these two putative virulence loci are shared byall other type I strains, indicating that the contribution of these genes to acute virulence iscommon to the entire lineage.
Other studies have indicated that additional genes may contribute to virulence in different
genetic backgrounds. A genetic cross originally performed between the non-virulent typesII and III strains (ave rise to at least one clone with enhanced virulencerelative to either parent (Importantly, the level of virulence exhibitedby this clone, while elevated from the parental types, is not equal to that of the type I strains. Because of the nature of this cross (both parental strains are non-virulent) it is not possibleto assign chromosomal loci controlling this trait with any confidence. However, a geneticbackcross followed by linkage analysis could be used to identify loci that contribute tovirulence in this system. L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Humans may become infected with T. gondii by ingesting oocysts shed into the environ-
ment by cats or by eating undercooked meat that harbors cysts of the parasite. The relativecontribution of each of these modes of infection to the overall burden of toxoplasmosisin humans is unknown. While infections with T. gondii only account for ∼1% of all foodborne infections, they comprise a large percentage of serious cases of disease leading tohospitalization and death (Specific outbreaks of toxoplasmosis havebeen reported in association with contaminated, unfiltered water supplies as typified by anoutbreak of British Columbia, Canada (Likewise, in highly endemicregions in Brazil of unfiltered water sources that werelikely contaminated by oocysts was found to be a risk factor for infection. Epidemiologystudies indicate that while contact with cats is not significant, exposure to soil is a risk fac-tor, indicating an important role for oocysts in transmission Moreover,eating undercooked meat is a strong risk factor for infection in pregnant women studied aspart of a European multicenter study (Numerous studies have highlightedthe prevalence of T. gondii in meat sources that are consumed as food and there are severaldocumented examples of transmission from ingestion of undercooked meat (Collectively, these studies highlight the importance of toxoplasmosis as a water orfood borne infection, although the relative proportions of each mode of transmission likelyvaries with locality and over time.
Serological methods for diagnosis of infection are well developed and are primarily based
on detecting IgM/IgA followed by conversion to predominantly IgG responses to parasiteantigens (The primary antigens of the acute infection are SAG1,a major tachyzoite surface protein, and a family of SAG-related antigens While members of this family are divergent and antigenicallydistinct from each other, each is well conserved with only minor variation seen betweenisolates (to the fact that most major antigensare highly conserved between strains, no system for serotyping has been established for T. gondii
Similar to the situation in mice, type I strains may be more virulent in humans based on
the observation that they are more prevalent in cases of human disease. Initial studies on theprevalence of T. gondii strains in animal and human infections indicated that type I strainswere over-represented in human toxoplasmosis compared to their distribution in animalsSince animals provide the source for most human infections,this pattern suggested that type I strains where either more infectious to humans or morelikely to result in severe disease and hence be isolated following clinical complications. Subsequent studies have confirmed this pattern in some populations (ut not in others These differences may reflectgeographic differences in strain prevalence, biases in strain collection due to either referralor isolation strategies, or underlying differences in patient populations. Further studies areneeded on a much broader scale in order to determine to what extent the genotype of theparasite contributes to the progression of disease in human toxoplasmosis.
Using a combination of DNA polymorphisms it is possible to unambiguously type all
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
provides a useful tool for molecular epidemiology studies. However, this approach relieson abundant material for accurate analysis. Consequently, it is not possible to type chronicinfections and hence we do not know the profile of strains that cause subclinical infec-tions. Despite strong antigenic similarity, several strain-specific antigenic differences havebeen defined in the murine model (). Recent advances in detecting antigenic differences that arestrain-specific indicate that it may be possible to type strains that cause infections basedon serology (This advance will provide an important denominator forestablishing the extent to which different strain types cause infection versus disease. 4. A recent origin for T. gondii
Initial attempts to define polymorphisms in T. gondii highlighted an unusually low het-
erozygosity in this genus using RFLP (r isoenzyme (markers. The sequence of antigen encoding genes from the different strain typesrevealed a simple dimorphic pattern for SAG2 SAG1 and GRA4 (When this pattern was examined on awider basis, it became clear that at each locus there exist just two distinct alleles, allowingfor minor variation due to mutation (This striking pattern led to theimportant conclusion that most of the current strains of T. gondii were derived from a singlegenetic cross between two parental strains (There are two extremely cu-rious features about this event. First, the two alleles themselves are quite similar, indicatingthat the mating took place between two very similar isolates. This may indicate an overalllow diversity in the population prior to this cross. Second, only three major lineages survivedthis cross despite the fact that recombination should have created many distinct mixturesof the two-allele system. What event could create and maintain such a bizarre pattern?
The striking clonality and evidence that most existing strains of T. gondii arose from two
closely related parental strains is at odds with the ancient nature of this phylum. This raisesimportant questions about the relative age of the genus Toxoplasma and how it is relatedto what are perceived as “closely related” members of the Apicomplexa. As mentionedpreviously, life cycle characters are not a reliable indication of common ancestry. Instead,molecular phylogenies are more likely to reflect historical relationships between T. gondiiand other tissue-cyst forming coccidians.
One of the advantages of molecular phylogenies is that they provide estimates of both
historical relatedness and relative age since a common ancestry. One of the most robustindexes for phylogenetic comparison is the rRNA encoding genes for the small subunit(SSU). SSU phylogenies have been used extensively to analyze the relationships withinthe Apicomplexa (In the absence of a fossil record, this provides a reliableand easily obtainable estimate of common ancestries for this group. Various estimates havebeen made for the average rate of nucleotide change within SSU genes for different taxa andusing an average value for this mutation rate, the divergences of the major groupings withinthe Apicomplexa have been estimated Using this approach,it has been estimated that coccidians diverged from the piroplasms and malarias some 800million years ago
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Fig. 1. Molecular phylogeny of T. gondii and related parasites. Ages were calculated using an estimate of theaverage mutation rate in the SSU sequences. Neighbor-joining phylogram based on aligned SSU sequences usingClustalW and PAUP 4.0. Accession number for SSU sequences are shown in the figure within parentheses.
Within the tissue-cysts forming coccidia, Toxoplasma is closely related to Neospora and
Hammondia, each of which contains several species. Using an estimate of the average rateof change for the SSU sequence, it was recently predicted that this group shares a commonancestry some 12 million years ago ((In contrast, the commonancestry with other tissue-cyst forming coccidians such as Sarcocystis spp. is ∼250 millionyears ago Interestingly, the divergence of T. gondii from Neospora and Hammondiaclosely correlates with the divergence of their respective definitive hosts Analysis of the internal transcribed spacer 1 (ITS-1) region of the rDNA genes indicatesHammondia is paraphyletic. Thus, while H. heydorni and N. caninum utilize the dog asdefinitive host, T. gondii and H. hammondia utilize the cat. Consequently, these taxa mayhave initially diverged based on their choice of definitive hosts before speciation led to thecurrent groupings.
The high degree of similarity between T. gondii strains suggests a much more recent
ancestry than would be predicted from the relationship of the genus Toxoplasma to otherclosely related parasites. The pattern of two alleles being shared between three commonlydistributed clonal lineages also suggests a recent genetic cross was responsible for givingrise to most of the existing strains of T. gondii. Consequently, we estimated the time since arecent common ancestry of the three predominant lineages (thatarose within both non-coding (introns) and coding regions of a number of unlinked geneswere analyzed among a group of different strains. Sequencing of these regions revealed theexpected dimorphic polymorphisms that define the clonal lineages When these shared polymorphisms were masked, only two unique mutations were observedin more than 40,000 bp of sequence from the clonal lineages. Age estimates based on aneutral mutation rate indicated that the three clonal lineages share a common ancestrywithin the last 10,000 years (
While the majority of T. gondii strains belong to one of the three predominant clonal
lineages, a few isolates have mixed genotypes (a result of recombination between the majortypes), and fewer still have rare genotypes that are highly polymorphic. These exotic strainshave been found in a variety of geographic locations and infect both humans and animalsWhat makes them exotic is that they containpolymorphisms that are not typical of the clonal lineages. When their age was estimated
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
based on these additional polymorphisms, it was found that these exotic strains originatedapproximately 1 million years ago (They clearly predate the origin of thethree predominant lineages, raising the possibility that they represent an ancestral state.
The extreme success of the three clonal lineages since their recent origin is remarkable.
They currently populate a wide range of mammalian and avian orders, which are separatedin evolutionary time by hundreds of millions of years. This pattern suggests that followinga recent origin, they rapidly expanded their host range, either replacing previously existingstrains of the parasite that were less fit, or colonizing new hosts for the first time. Whattrait(s) were they endowed with that led to this remarkable success?
The key life cycle feature that distinguishes T. gondii from its close relatives is the lack
of reliance on the definitive host for transmission (Because tissue cysts of T. gondiiare orally infectious for other intermediate hosts, transmission can occur without a need forinfection of the cat. This ability is likely important for maintaining clonality. However, itsreal significance is that it likely arose recently and may have lead to the extreme successof the predominant clonal lineages. This model is supported by the fact that lack of directoral infectivity is an ancestral trait shared by all close relatives of T. gondii. Furthermore,while the clonal lineages of T. gondii possess oral infectivity, some of the exotic lineages aredeficient or completely lacking in this trait (Collectively these data led to
Fig. 2. Life cycle of typical tissue cysts forming coccidian versus T. gondii. The restricted two-host life cycleof tissue-cyst forming coccidians involves obligatory passage through the definitive host where the sexual cycleoccurs (left). Ingestion of animal tissues harboring cysts only leads to infection in the definitive host (those ableto support development of sexual phases). Within the definitive host, development occurs in the gut and oocystsare shed into the environment where they are picked up by intermediate hosts. Transmission of T. gondii followsa similar two-host cycle, in which members of the cat family serve as the definitive host although many speciesserve as intermediate hosts. However transmission of T. gondii also occurs between intermediate hosts via directoral infection (right). Here ingestion of tissue cysts leads to infection even in other intermediate hosts. Infectionscan be spread by cannibalism, omnivorous feeding, carnivorous feeding, and scavenging, thus greatly expandingthe infection of new hosts. L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
the model that oral infectivity arose as the result of a recent cross between two highly similarparental types. Presumably the mutations that gave rise to oral infectivity existed separatelyin the parental strains, where they likely did not provide any selective advantage. Only whenthese separate mutations (alleles) came together via reassortment or recombination did oralinfectivity result. This event was evidently very rare as only three distinct lineages survived,suggesting the trait is multigenic.
Acquisition of direct oral infectivity can be defined as a “key adaptation” as it would have
enabled these strains to spread more rapidly through the food chain via direct transmission. Instead of having to infect the cat in each cycle, scavenging or carnivorous feeding wouldlead to infection of new intermediate hosts. In bypassing the need for the sexual cycle, directoral infectivity also led to a rapid fixing of the entire genome via a “hitch hiking effect”This mechanism is likely responsible for creating theextreme linkage disequilibrium of the present day clonal lineages.
Importantly, the exotic strains express oral infectivity to a variable extent, some being
completely non-infectious while others are fully infectious (While thisseems at odds with the above model, a closer look at their genomes reveals why this re-sult is entirely expected. The exotic strains, while clearly ancestral in many respects alsoshow evidence of the dimorphic allele patterns that typifies the clonal lineages (This strongly suggests that they have introgressed (interbred) with the clonal lin-eages through subsequent crosses, and consequently acquired variable expression of directoral infectivity. True ancestral lineages may be extremely rare as they are restricted to anobligate two-host transmission cycle.
The time frame for the origin of T. gondii strains with direct oral infectivity coincides
closely with the domestication of companion and agricultural animals (Thisevent likely led to exposure of new hosts to parasites they had not previously directly en-countered and may have provided strong selective pressure for the emergence of new modesof transmission. For example, practices such as feeding offal from agricultural animals todogs and cats would likely expose them to parasites they would not naturally encounterin the wild. Because cats are often semi-domesticated in farm environments, they are alsoexposed to parasites found in sylvatic cycles. Furthermore, the housing of agricultural ani-mals in close quarters could assure the continued passage of this unusual mixture of strainsby an fecal–oral route through contamination of feed, water, or pastures. 5. Conclusions and perspectives
While complex traits such as life cycle features have traditionally been used for inferring
phylogenetic relationships, closer inspection reveals that they are not well suited for suchanalyses. Rather, such complex features may be highly unstable given the possibilitiesfor expansion into new niches that arise when they are altered. It is counter-intuitive thatcomplex traits such as host range and infectivity would be easily changed and this challengesthe idea that host parasite relationships are highly evolved and stable relationships. Atpresent nothing is known about the molecular mechanisms for bringing about such changes. However, they presumably arise through the accumulation of mutations that can rapidly andprofoundly alter fitness given the right combination of opportunity and selection. L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Toxoplasma provides a model example of how complex life cycle traits can change
rapidly over short periods of time. Changes in exposure patterns that occurred with thedomestication of animals likely played a crucial role in creating opportunity for this eventby providing new niches for exploitation. In T. gondii, sexual recombination provided thecritical reshuffling of the genes that led to direct oral infectivity. This trait likely contributesto the wide spread success of T. gondii, which is one of the world’s most common para-sitic protozoan infections of animals and humans. Because oral infectivity resulted in thesimultaneous bypassing of sexual recombination, the genotypes that acquired this new traithave remained essentially fixed since their origin. Other large-scale changes in life cyclesmay have occurred without being accompanied by genetic fixation, making them harderto detect. Consequently, dramatic changes in transmission dynamics, host range, and de-velopment among parasitic life styles may be more widespread in nature than previouslyassumed. Acknowledgements
Funding from the USDA, NIH, and the Burroughs Wellcome Fund has supported the
work in my laboratory. I am grateful to current and past members of my laboratory whohave conducted much of the work described here as well as colleagues and collaboratorswhom have provided valuable assistance and critical input. References
Aikawa, M., Sterling, C.R., 1974. Intracellular Parasitic Protozoa. Academic Press, New York. Ajzernberg, D., Cogné, N., Paris, L., Bessieres, M.H., Thulliez, P., Fillisetti, D., Pelloux, H., Marty, P., Dardé,
M.L., 2002. Genotype of 86 Toxoplasma gondii isolates associated with human congenital toxoplasmosis andcorrelation with clinical findings. J. Infect. Dis. 186, 684–689.
Anderson, T.J., Haubold, B., Williams, J.T., Estrada-Franco, J.G., Richardson, L., Mollinedo, R., Bockarie, M.,
Mokili, J., Mharakurwa, S., French, N., Whitworth, J., Velez, I.D., Brockman, A.H., Nosten, F., Ferreira, M.U.,Day, K.P., 2000. Microsatellite markers reveal a spectrum of population structures in the malaria parasitePlasmodium falciparum. Mol. Biol. Evol. 17, 1467–1482.
Bahia-Oliveira, L.M., Jones, J.L., Azevedo-Silva, J., Alves, C.C., Orefice, F., Addiss, D.G., 2003. Highly endemic,
waterborne toxoplasmosis in north Rio de Janeiro state, Brazil. Emerg. Infect. Dis. 9, 55–62.
Baldauf, S.L., Roger, A.J., Wenk-Siefert, I., Doolittle, W.F., 2000. A kingdom-level phylogeny of eukaryotes based
on combined protein data. Science 290, 972–977.
Barta, J.R., 1989. Phylogenetic analysis of the class sporozoea (Phylum Apicomplexa Levine 1970): evidence for
the independent evolution of heteroxenous life cycles. J. Parasitol. 75, 195–206.
Barta, J.R., 2001. Molecular approaches for inferring evolutionary relationships among protistan parasites. Vet.
Barta, J.R., Martin, D.S., Carreno, R.A., Siddall, M.E., Profous-Juchelkat, H., Hozza, M., Powles, M.A.,
Sundermann, C., 2001. Molecular phylogeny of the other tissue coccidia: Lankesterella and Caryospora. J. Parasitol. 87, 121–127.
Bohne, W., Gross, U., Heesemann, J., 1993. Differentiation between mouse-virulent and avirulent strains of
Toxoplasma gondii by a monoclonal antibody recognizing a 27-kilodalton antigen. J. Clin. Microbiol. 31,1641–1643.
Boothroyd, J.C., Hehl, A., Knoll, L.J., Manger, I.D., 1997. The surface of Toxoplasma: more and less. Int. J. L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Bowie, W.R., King, A.S., Werker, D.H., Issac-Renton, J.L., Eng, S.B., Marion, S.A., 1997. Outbreak of
toxoplasmosis associated with municipal drinking water. The BC Toxoplasma investigation team. Lancet 19,173–177.
Bulow, R., Boothroyd, J.C., 1991. Protection of mice from fatal Toxoplasma gondii infection by immunization
with p30 antigen in liposomes. J. Immunol. 147, 3496–3500.
Carreno, R.A., Martin, D.S., Barta, J.R., 1999. Cryptosporidium is more closely related to the gregarines than to
coccidia as shown by phylogeneyic analysis of apicomplexan parasites inferred using small-subunit ribosomalRNA gene sequences. Parasitol. Res. 85, 899–904.
Conway, D.J., Roper, C., Oduola, A.M., Armot, D.E., Kremsner, P., Grobusch, M.P., Curtis, C.F., Greenwood,
B.M., 1999. High recombination rate in populations of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA96, 4506–4511.
Cook, A.J., Gilbert, R.E., Buffolano, W., Zufferey, J., Petersen, E., Jenum, P.A., Foulon, W., Semprini, A.E.,
Dunn, D.T., 2000. Sources of toxoplasma infection in pregnant women: European multicentre case-controlstudy. European Research Network on Congenital Toxoplasmosis. Br. Med. J. 321, 142–147 (see comments).
Dardé, M.L., Bouteille, B., Pestre-Alexandre, M., 1988. Isoenzyme characterization of seven strains of Toxoplasmagondii by isoelectric focusing in polyacrylamide gels. Am. J. Trop. Med. Hyg. 39, 551–558.
Dardé, M.L., Bouteille, B., Pestre-Alexandre, M., 1992. Isoenzyme analysis of 35 Toxoplasma gondii isolates and
the biological and epidemiological implications. J. Parasitol. 78, 786–794.
Dubey, J.P., 1977. Toxoplasma, Hammondia, Besniotia, Sarcocystis, and other tissue cyst-forming coccidia of man
and animals. In: Kreier, J.P. (Ed.), Parasitic Protozoa. Academic Press, New York, pp. 101–237.
Dubey, J.P., Beattie, C.P., 1988. Toxoplasmosis of Animals and Man. CRC Press, Boca Raton, FL. Dubey, J.P., Lindsay, D.S., 1996. A review of Neospora caninum and neosporosis. Vet. Parasitol. 67, 1–59. Escalante, A.A., Ayala, F.J., 1995. Evolutionary origin of Plasmodium and other Apicomplexa based on rRNA
genes. Proc. Natl. Acad. Sci. USA 92, 5793.
Falkow, S., 1988. Molecular Kock’s postulates applied to microbial pathogenicity. Rev. Infect. Dis. 10, S274–S276. Fazaeli, A., Carter, P.E., Dardé, M.L., Pennington, T.H., 2000. Molecular typing of Toxoplasma gondii strains by
GRA6 gene sequence analysis. Int. J. Parasitol. 30, 637–642.
Ferguson, D.J.P., Hutchison, W.M., 1981. Comparison of the development of avirulent and virulent strains of
Toxoplasma gondii in the peritoneal exudate of mice. Ann. Trop. Med. Parasitol. 75, 539–546.
Fuentes, I., Rubio, J.M., Ram´ırez, C., Alvar, J., 2001. Genotypic characterization of Toxoplasma gondii strains
associated with human toxoplasmosis in Spain: direct analysis from clinical samples. J. Clin. Microbiol. 39,1566–1570.
Gajadhar, A.A., Marquardt, W.C., Hall, R., Gunderson, J., Ariztia-Carmona, E.V., Sogin, M.L., 1991. Ribosomal
RNA sequences of Sarcocystis muris, Theileria annulata and Crypthecodinium cohnii reveal evolutionaryrelationships among apicomplexans, dinoflagelates, and ciliates. Mol. Biochem. Parasitol. 45, 147–154.
Gavrilescu, L.C., Denkers, E.Y., 2001. IFN-␥ overproduction and high level apoptosis are associated with high
but not low virulence Toxoplasma gondii infection. J. Immunol. 167, 902–909.
Grigg, M.E., Bonnefoy, S., Hehl, A.B., Suzuki, Y., Boothroyd, J.C., 2001a. Success and virulence in Toxoplasma
as the result of sexual recombination between two distinct ancestries. Science 294, 161–165.
Grigg, M.E., Ganatra, J., Boothroyd, J.C., Margolis, T.P., 2001b. Unusual abundance of atypical strains associated
with human ocular toxoplasmosis. J. Infect. Dis. 184.
Gross, U., Muller, W.A., Knapp, S., Heesemann, J., 1991. Identification of a virulence-associated antigen of
Toxoplasma gondii by use of a mouse monoclonal antibody. Infect. Immun. 59, 4511–4516.
Hall, S., Ryan, M., Buxton, D., 2001. The epidemiology of toxoplasma infection. In: Joynson, D.H.M., Wreghitt,
T.G. (Eds.), Toxoplasmosis: A Comprehensive Clinical Guide. Cambridge University Press, Cambridge,pp. 58–124.
Handman, E., Remington, J.S., 1980. Antibody responses to Toxoplasma antigens in mice infected with strains of
different virulence. Infect. Immun. 29, 215–220.
Hole, F., 1994. Origins of Agriculture. Cambridge University Press, Cambridge. Howe, D.K., Sibley, L.D., 1995. Toxoplasma gondii comprises three clonal lineages: correlation of parasite
genotype with human disease. J. Infect. Dis. 172, 1561–1566.
Howe, D.K., Summers, B.C., Sibley, L.D., 1996. Acute virulence in mice is associated with markers on chromosomeVIII in Toxoplasma gondii. Infect. Immun. 64, 5193–5198. L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Howe, D.K., Honoré, S., Derouin, F., Sibley, L.D., 1997. Determination of genotypes of Toxoplasma gondii strains
isolated from patients with toxoplasmosis. J. Clin. Microbiol. 35, 1411–1414.
Johnson, J., Suzuki, Y., Mack, D., Mui, E.J., Estes, R., David, C., Skamane, E., Forman, J., McLeod, R., 2002.
Genetic analysis of influences on survival following Toxoplasma gondii infection. Int. J. Parasitol. 32, 179–185.
Joynson, D.H.M., Guy, E.C., 2001. Laboratory diagnosis of toxoplasma infection. In: Joynson, D.H.M., Wreghitt,
T.G. (Eds.), Toxoplasmosis: A Comprehensive Clinical Guide. Cambridge University Press, Cambridge,pp. 296–318.
Kempf, M.C., Cesbron-Delauw, M.F., Deslee, D., Gross, U., Herrmann, T., Sutton, P., 1999. Different
manifestations of Toxoplasma gondii infection in F344 and LEW rats. Med. Microbiol. Immunol. (Berl) 187,137–142.
Kissinger, J.C., Collins, W.E., Li, J., McCutchan, T.F., 1998. Plasmodium inui is not closely related to other quartan
plasmodium species. J. Parasitol. 84, 278–282.
Kong, J.T., Grigg, M.E., Uyetake, L., Parmley, S., Boothroyd, J.C., 2003. Serotyping of Toxoplasma gondii
infections in humans using synthetic peptides. J. Infect. Dis. 187, 1484–1495.
Lander, E.S., Botstein, D., 1989. Mapping Mendelian factors underlying quantitative traits using RFLP linkage
Lander, E., Kruglyak, L., 1995. Genetic dissection of complex traits: guidelines for interpreting and reporting
linkage results. Nat. Genet 11, 241–247.
Levine, N.D., 1988. The Protozoan Phylum Apicomplexa. CRC Press, Boca Raton, FL. Liesenfeld, O., Kosek, J., Remington, J.S., Suzuki, Y., 1996. Association of CD4+ T cell-dependent,
interferon-␥-mediated necrosis of the small intestine with genetic susceptibility of mice to peroral infectionwith Toxoplasma gondii. J. Exp. Med. 184, 597–607.
Liesenfeld, O., Kang, H., Park, D., Nguyen, T.A., Parkhe, C.V., Watanabe, H., Abo, T., Sher, A., Remington,
J.S., Suzuki, Y., 1999. TNF␣, nitric oxide and IFN-␥ are all critical for development of necrosis in the smallintestine and early mortality in genetically susceptible mice infected perorally with Toxoplasma gondii. ParasiteImmunol. 21, 365–376.
Manger, I., Hehl, A.B., Boothroyd, J.C., 1998. The surface of Toxoplasma tachyzoites is dominated by a family
of glycosylphosphatidylinositol-anchored antigens related to SAG1. Infect. Immun. 66, 2237–2244.
Maynard Smith, J., Haigh, J., 1974. Genet. Res. 23, 23–35. McLeod, R., Estes, R.G., Mack, D.G., Cohen, H., 1984. Immune response of mice to ingested Toxoplasma gondii:
a model of toxoplasma infection acquired by ingestion. J. Infect. Dis. 149, 234–244.
Mead, P.S., Slutsker, L., Dietz, V., McCaig, L.F., Bresee, J.S., Shapiro, C., Griffin, P.M., Tauxe, R.V., 1999.
Food-related illness and death in the United States. Emerg. Infect. Dis. 5, 607–625.
Meisel, R., Stachelhaus, S., Mévélec, M., Reichmann, G., Dubremetz, J.F., Fischer, H., 1996. Identification of two
alleles in the GRA4 locus of Toxoplasma gondii determining a differential epitope which allows discriminationof type I versus types II and III strains. Mol. Biochem. Parasitol. 81, 259–263.
Mordue, D.G., Monroy, F., La Regina, M., Dinarello, C.A., Sibley, L.D., 2001. Acute toxoplasmosis leads to lethal
overproduction of Th1 cytokines. J. Immunol. 167, 4574–4584.
Parmley, S.F., Gross, U., Sucharczuk, A., Windeck, T., Sgarlato, G.D., Remington, J.S., 1994. Two alleles of the
gene encoding surface antigen P22 in 25 strains of Toxoplasma gondii. J. Parasitol. 80, 293–301.
Perkins, S.L., Schall, J.J., 2002. A molecular phylogeny of malarial parasites recovered from cytochrome b gene
sequences. J. Parasitol. 88, 972–978.
Pfefferkorn, E.R., Kasper, L.H., 1983. Toxoplasma gondii: genetic crosses reveal phenotypic suppression of
hydroxyurea resistance by fluorodeoxyuridine resistance. Exp. Parasitol. 55, 207–218.
Roos, D.S., Donald, R.G.K., Morrissette, N.S., Moulton, A.L., 1994. Molecular tools for genetic dissection of the
protozoan parasite Toxoplasma gondii. Meth. Cell Biol. 45, 28–61.
Sabin, A.B., 1941. Toxoplasmic encephalitis in children. J. Am. Med. Assoc. 116, 801–807. Sibley, L.D., Boothroyd, J.C., 1992. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature
Sibley, L.D., LeBlanc, A.J., Pfefferkorn, E.R., Boothroyd, J.C., 1992. Generation of a restriction fragment length
polymorphism linkage map for Toxoplasma gondii. Genetics 132, 1003–1015.
Sogin, M.L., Silberman, J.D., 1998. Evolution of the protists and protistan parasites from the perspective of
molecular systematics. Int. J. Parasitol. 28, 11–20. L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Su, C., Howe, D.K., Dubey, J.P., Ajioka, J.W., Sibley, L.D., 2002. Identification of quantitative trait loci controlling
acute virulence in Toxoplasma gondii. Proc. Natl. Acad. Sci. USA 99, 10753–10758.
Su, C.L., Evans, D., Cole, R.H., Kissinger, J.C., Ajioka, J.W., Sibley, L.D., 2003. Enhanced oral transmission
acquired through recent recombination led to widespread expansion of Toxoplasma. Science 299, 414–416.
Suzuki, Y., Joh, K., Orellana, M.A., Conley, F.K., Remington, J.S., 1991. A gene(s) within the H-2D region
determines the development of toxoplasmic encephalitis in mice. Immunology 74, 732–739.
Suzuki, Y., Joh, K., Kwon, O.C., Conley, F.K., Remington, J., 1994. MHC class I gene(s) in the D/L region but not
the TNF-␣ gene determines development of toxoplasmic encephalitis in mice. J. Immunol. 153, 4649–4654.
Tibayrenc, M., Kjellberg, F., Ayala, F.J., 1990. A clonal theory of parasitic protozoa: the population structures of
Entamoeba, Giardia, Leishmania, Naegleria, Plasmodium, Trichamonas, and Trypanosoma and their medicaland taxonomic consequences. Proc. Natl. Acad. Sci. USA 87, 2414–2418.
Weiss, L.M., Udem, S.A., Tanowitz, H., Wittner, M., 1987. Western blot analysis of the antibody response of
patients with AIDS and toxopalsmic encephalitis: antigenic diversity among Toxoplasma strains. J. Infect. Dis. 157, 7–13.
Wray, G.A., Leviton, J.S., Shapiro, L.H., 1996. Molecular evidence for deep precambrian divergences among
Zenner, L., Darcy, F., Capron, A., Cesbron-Delauw, M.F., 1998. Toxoplasma gondii: kinetics of the dissemination
in the host tissues during the acute phase of infection in rats and mice. Exp. Parasitol. 90, 86–94.
Zenner, L., Foulet, A., Caudrelier, Y., Darcy, F., Gosselin, B., Capron, A., Cesbron-Delauw, M.F., 1999. Infection
with Toxoplasma gondii RH and Prugniaud strains in mice, rats and nude rats: kinetics of infection in bloodand tissues related to pathology in acute and chronic infection. Pathol. Res. Pract. 195, 475–485.
Sexual Dysfunction Association www.sda.uk.net Email: [email protected] VIAGRA VIAGRA (sildenafil) is the first oral treatment for erectile dysfunction (impotence) to be licenced in the UK. This fact sheet provides basic information on VIAGRA; for more detailed information you should consult your doctor. What is VIAGRA? VIAGRA belongs to a group of medicines called phosphodieste
TO: Columbus Area EMS District Board Members FROM: Werner Biedermann DATE: July 10, 2013 SUBJECT: June 2013 Monthly Report Lifestar responded to 54 emergency 911 calls in June. We also performed a paramedic intercept for Randolph EMS and one fire standby for Fall River Fire Department. Below is the call breakdown by municipality. Columbus Contract Run Statistics for 2013 JA


 Veterinary Parasitology 115 (2003) 185–198
Department of Molecular Microbiology, Washington University School of Medicine,
660 S Euclid Box 8230, St. Louis, MO 63110, USA
Abstract
Veterinary Parasitology 115 (2003) 185–198
Department of Molecular Microbiology, Washington University School of Medicine,
660 S Euclid Box 8230, St. Louis, MO 63110, USA
Abstract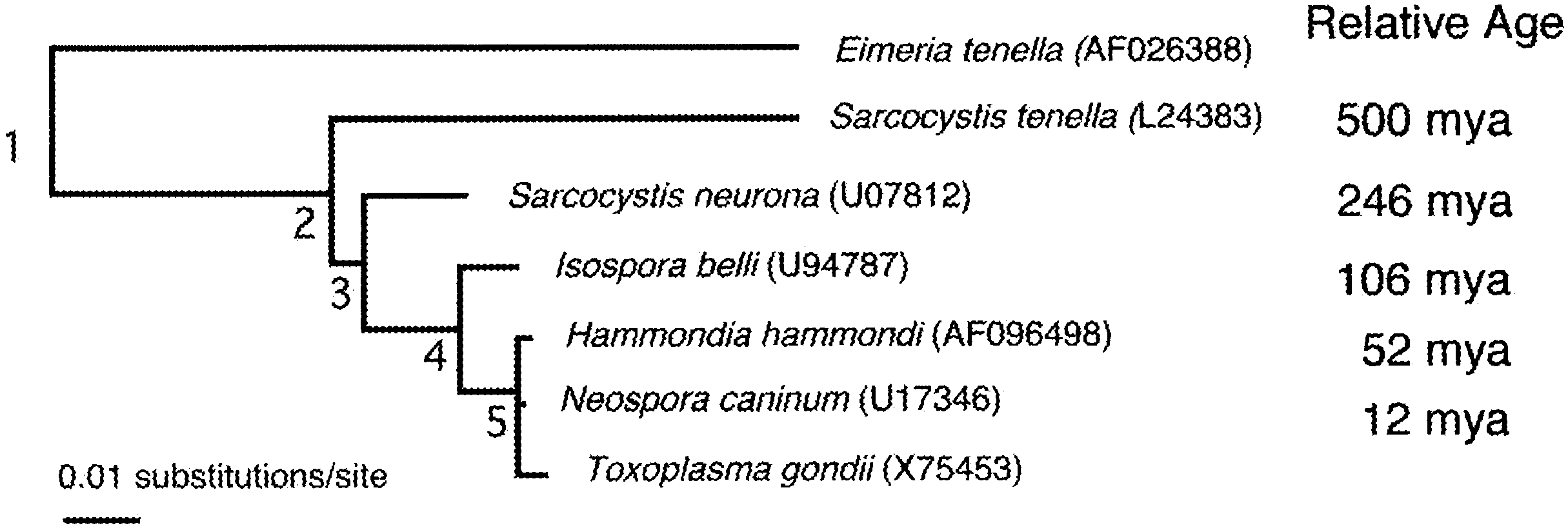 L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Fig. 1. Molecular phylogeny of T. gondii and related parasites. Ages were calculated using an estimate of theaverage mutation rate in the SSU sequences. Neighbor-joining phylogram based on aligned SSU sequences usingClustalW and PAUP 4.0. Accession number for SSU sequences are shown in the figure within parentheses.
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
Fig. 1. Molecular phylogeny of T. gondii and related parasites. Ages were calculated using an estimate of theaverage mutation rate in the SSU sequences. Neighbor-joining phylogram based on aligned SSU sequences usingClustalW and PAUP 4.0. Accession number for SSU sequences are shown in the figure within parentheses.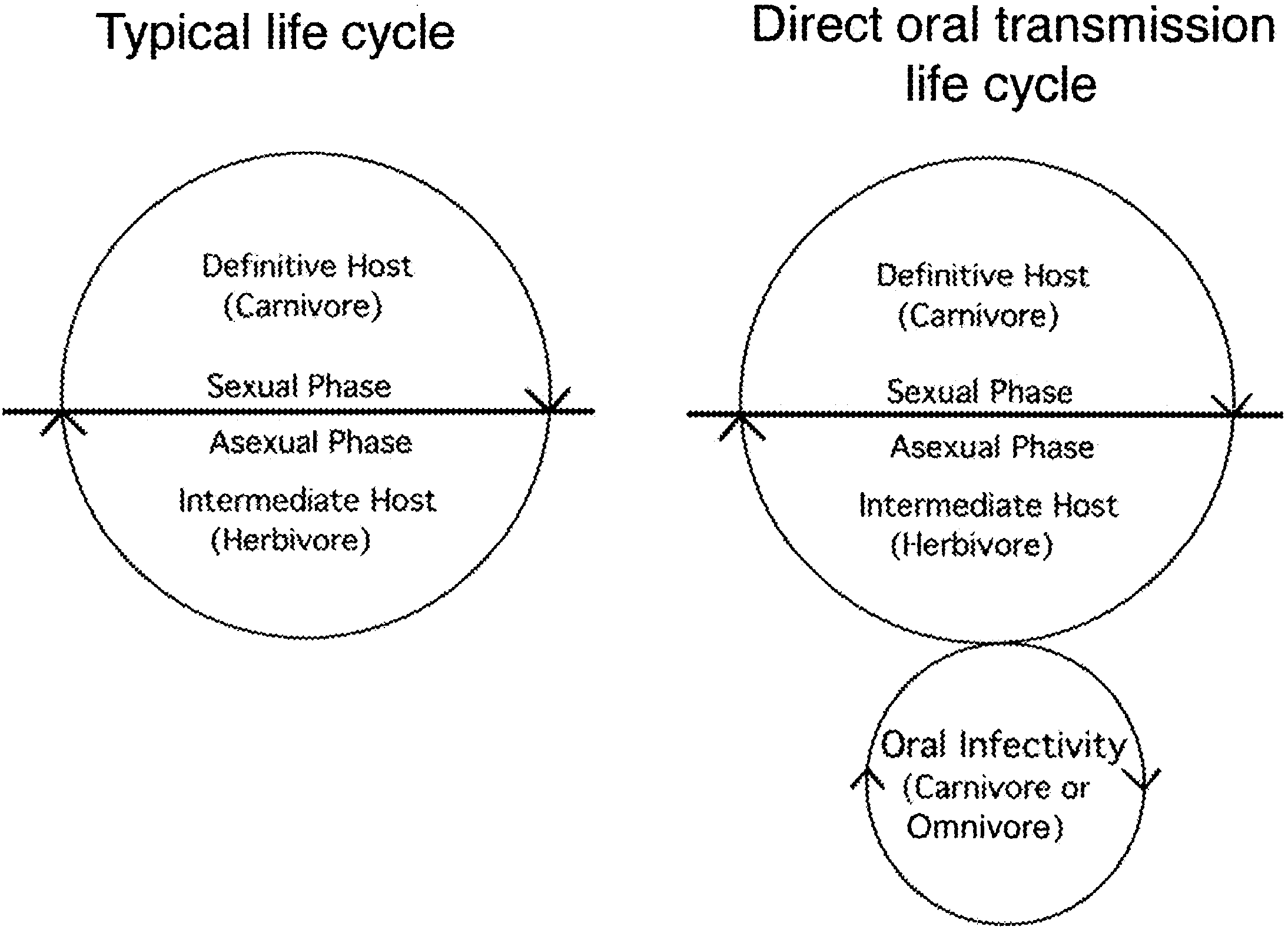 L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
based on these additional polymorphisms, it was found that these exotic strains originatedapproximately 1 million years ago (They clearly predate the origin of thethree predominant lineages, raising the possibility that they represent an ancestral state.
L. David Sibley / Veterinary Parasitology 115 (2003) 185–198
based on these additional polymorphisms, it was found that these exotic strains originatedapproximately 1 million years ago (They clearly predate the origin of thethree predominant lineages, raising the possibility that they represent an ancestral state.