The Journal of Experimental Biology 205, 297–306 (2002)
Printed in Great Britain The Company of Biologists Limited 2002JEB3755
Trimethylamine oxide accumulation in marine animals: relationship to acylglycerol NIEHS Marine and Freshwater Biomedical Sciences Center, Rosenstiel School of Marine and Atmospheric Science,
*Present address: Monterey Bay Aquarium Research Institute, Moss Landing, CA 95039, USA (e-mail: [email protected])
Trimethylamine oxide (TMAO) is a common and as diacylglycerol ethers and triacylglycerols. TMAO is compatible osmolyte in muscle tissues of marine synthesized from the trimethylammonium moiety of organisms that is often credited with counteracting choline, thus released, and is retained as a compatible protein-destabilizing forces. However, the origin and solute in concentrations reflecting the amount of lipid synthetic pathways of TMAO are actively debated. Here, stored in the body. A variation on this theme is proposed we examine the distribution of TMAO in marine animals for sharks. and report a correlation between TMAO and acylglycerol storage. We put forward the hypothesis that TMAO is derived, at least in part, from the hydrolysis of
Key words: trimethylamine oxide, choline, phosphatidylcholine,
phosphatidylcholine, endogenous or dietary, for storage
lipid, cephalopod, buoyancy, deep sea, urea, solute. Introduction
Methylamine compounds, particularly trimethylamine oxide
a review, see Ballantyne, 1997), the counteracting solute
(TMAO), are compatible osmolytes that commonly occur in
hypothesis has received wide support (Wang and Bolen, 1997;
tissues of marine organisms (Yancey et al., 1982). Their
Yancey et al., 1982; Yancey and Somero, 1980; Yancey and
concentrations vary extensively, however, among habitats and
species and even with season and ontogeny within species (for
TMAO may also counteract the effects of hydrostatic
a review, see Hebard et al., 1982). There are numerous
pressure on enzyme function in deep-sea animals. Yancey and
hypotheses attempting to account for the distribution of
colleagues (Gillett et al., 1997; Kelly and Yancey, 1999) have
methylamines. High TMAO levels in polar fishes are thought
demonstrated a correlation between TMAO concentration and
to increase osmotic concentration, thus depressing the freezing
capture depth in a variety of organisms. They have also shown
point of the body fluids (Raymond and DeVries, 1998;
that 250 mmol l–1 TMAO in vitro is able to counteract the loss
Raymond, 1994). Sanders and Childress (1988) and Withers et
of activity in some enzymes that results from high hydrostatic
al. (1994) point out that trimethylamine (TMA) and TMAO,
pressure (Yancey and Siebenaller, 1999; Yancey et al., 2001).
as a result of their large positive partial molal volumes, impart
Other methylamines are known to counteract the effects of
considerable lift to counteract sinking in some pelagic marine
ammonia toxicity (Kloiber et al., 1988; Minana et al., 1996),
animals. TMAO is best known, however, as a ‘counteracting
salt (Dragolovich, 1994) and temperature stress (Krall et al.,
solute’ that protects proteins against various destabilizing
1989; Nishiguchi and Somero, 1992) on protein function in
a variety of organisms. Although TMAO may serve the
In elasmobranchs, for example, TMAO may counteract the
functions attributed to it (i.e. it may be adaptive), it is not
toxic effects of urea on proteins (Somero, 1986; Yancey and
necessarily synthesized or accumulated as a specific adaptation
Somero, 1980). Because they are iso-osmotic with sea water,
to any of the stresses mentioned above. The source of TMAO
sharks retain large quantities of urea in their fluids as an
in marine animals and the time course of its accumulation
osmolyte. Urea is highly perturbing to enzyme systems.
are not well characterized. Here, we present preliminary
TMAO has a demonstrated ability to counteract the perturbing
measurements of TMAO concentrations in a number of
effects of urea on enzyme activity when accumulated in a 2:1
cephalopod species and review the distribution, biosynthesis
ratio with urea (Somero, 1986). Although some enzymes seem
and metabolism of methylamines in marine animals to assess
to have an evolved tolerance of urea regardless of TMAO (for
Methylamine synthesis
exclusively in the gut. However, the mechanism for conversion
TMAO, like most methylamines, is derived from the
of choline to TMA other than by microbial oxidation is not
trimethylammonium group of choline. Dietary choline may be
oxidized to trimethylamine by bacteria in the gut of marine
Furthermore, fasted dogfish maintained stable TMAO
animals. The accumulation of TMA in rotting fish as a result of
concentrations over 41 days (Cohen et al., 1958). This result
bacterial degradation of choline, as well as the reduction of
may be partly due to the active reabsorption of TMAO in the
TMAO to TMA, is responsible for their characteristic ‘fishy’
dogfish kidney (Cohen et al., 1958; Goldstein et al., 1967).
odor. TMA as a spoilage index for commercial fishes has been
Some sharks retain TMAO quite effectively, but substantial
extensively discussed (Hebard et al., 1982; Sotelo and Rehbein,
loss does occur such that maintenance of TMAO levels over
2000). TMA is highly toxic (Marzo and Curti, 1997; Anthoni
41 days seems unlikely without an endogenous source.
et al., 1991a,b) and so is, with at least one notable exception
Goldstein and Palatt (1974) found TMAO turnover rates of
(Sanders and Childress, 1988), oxygenated within living
4–14 % per day in four different elasmobranch species, but
animals to form TMAO (Fig. 1). This generally occurs within
excretion rates of less than 1 % have been reported in some
the digestive gland or liver via a monoxygenase enzyme,
fishes (Agustsson and Strøm, 1981). Diets rich in TMA and
trimethylamine oxidase (Tmase) (Hebard et al., 1982). TMAO
choline have produced higher levels of TMAO in the muscles
is then either transported to the tissues for accumulation as a
of some fish species but not in others (Goldstein et al., 1967;
compatible or ‘counteracting’ osmolyte or, more commonly,
Agustsson and Strom, 1981). An endogenous source is
excreted. In humans, a mutation of the flavin-containing mono-
suspected in many species, especially those that accumulate
oxygenase gene (FMO3) causes trimethylaminuria, a condition
wherein individuals excrete TMA, rather than TMAO, along
Endogenous choline supply is believed to limit the
with a fishy body odor in urine, sweat, breath and other bodily
accumulation of glycine betaine (betaine), another common
excretions (Dolphin et al., 1997; Treacy et al., 1998).
methylamine osmolyte, in euryhaline oysters (Pierce et al.,
There are, however, conflicting reports regarding the
1997). Oyster mitochondria take up choline and convert it, via
endogenous or exogenous (dietary) originof TMAO in animals. Early reports
suggested that invertebrates lacked Tmaseactivity (Baker et al., 1963). However,
phytoplankton production (Strøm, 1979).
Trimethylamine oxidase activity has beendetected in many, but not all, fishes
(Baker et al., 1963). Among polar fishes,
concentrations of TMAO than did speciesin which Tmase was detected (Raymondand DeVries, 1998). Goldstein et al.
TMAO in dogfish (Squalus acanthius)
Tmase activity (Baker et al., 1963). They
suggested that TMAO must beaccumulated from the diet. However,
detected Tmase activity in dogfish as wellas in the silky shark Carcharhinusfalciformis. Nurse sharks also possess
Fig. 1. Diagram showing the possible pathways for trimethylamine (TMA) and
Tmase activity and are able to synthesize
trimethylamine oxide (TMAO) production (only abbreviated pathways are drawn).
TMAO directly from choline in vivo
(A) Trimethylalkylammonium compounds (e.g. choline) are degraded to TMA by intestinal
microbes (Hebard et al. 1992). (B) Choline is taken up by mitochondria in some animals
(Pierce et al., 1997) and converted to glycine betaine via betaine aldehyde. Betaine may
subsequently be converted to TMA (Ballantyne, 1997). In both cases (A and B), TMA is
oxidized by trimethylamine oxygenase to TMAO. Trimethylamine oxide in marine animals
betaine aldehyde, to betaine (Fig. 1B). The enzymes involved
ethanolamine (Hanson and Rhodes, 1983; Summers and
in this transformation are well characterized (Dragolovich,
Weretilnyk, 1993; Weretilnyk et al., 1995). Radioactively
1994). Betaine could, if similarly produced in other species,
labeled ethanolamine in spinach is recovered primarily as
subsequently be converted to TMA and TMAO (Fig. 1B).
betaine. In contrast, labeled ethanolamine in wheat and barley
Some studies suggest that TMAO is a better counteracting
is recovered primarily in phospholipid (Hitz et al., 1981;
McDonnell and Wyn Jones, 1988) (Fig. 2).
Some plants produce methylamines, particularly betaine,
Wheat does appear to accumulate betaine, but only through
reportedly for osmoregulation and protection against drought
phosphatidylcholine hydrolysis (Fig. 2B). In wheat and barley
and salt-stress. A tremendous amount of research has been
leaves, phosphatidylcholine is hydrolyzed to diacylglycerol and
directed towards elucidating the pathways of betaine
choline. A similar pathway is suspected for oysters and
accumulation in plants in the hope of conferring drought-
horseshoe crabs, which convert choline to betaine as well (Pierce
resistance to important commercial crops such as tobacco
et al., 1997). Diacylglycerol, produced via phosphatidylcholine
(Nuccio et al., 1998). Plant research may, thus, provide insight
hydrolysis in wheat plants, is subsequently converted to
into the pathways and mechanisms of methylamine synthesis
monogalactosyldiacylglycerol, an essential glycolipid in
and storage in marine animals. Several pathways for the
photosynthetic membranes. Free choline is converted to betaine
production of choline in plants have been identified. Drought-
(McDonnell and Wyn Jones, 1988). In drought-resistant
resistant plants, such as chenopods (e.g. spinach and sugar
chenopods, betaine accumulation does not depend on
beet), are adapted for direct production of choline from
phosphatidylcholine hydrolysis, a fact that may result in their
Lipid storagePhotosynthetic membrane glycolipidSecond messengerFatty acid oxidation
Fig. 2. (A) Diagram of the glycerolphosphate pathway and phosphatidylcholine synthesis as found in both animals and plants. The bold arrowsindicate hypothesized pathways resulting in simultaneous accumulation of acylglycerols and trimethylamine oxide (TMAO) (only abbreviatedpathways are drawn). Diacylglycerol (DAG), formed from the derivative glycerol-3-phosphate (DAG), can be shunted towards eitheracylglycerol (lipid) storage (e.g. triacylglycerol) or phosphatidylcholine (PtdCho) synthesis. The final step in the phosphatidylcholine pathwayis reversible. The back reaction may occur to a significant extent so that diacylglycerol is formed from phosphatidylcholine and subsequentlystored for seasonal or reproductive energy reserves (see Gur and Harwood, 1991). Choline, thus released, is oxidized to TMAO and eitherstored or excreted. (B) In some plants, PtdCho is produced by methylation of ethanolamine and may subsequently be hydrolysed for release ofDAG and choline. DAG in plants is important during growth for the formation of photosynthetic membrane glycolipids. Choline, thus released,is oxidized to glycine betaine (Hitz et al. 1981). (C) In chenopods (e.g. spinach and sugarbeet), choline is synthesized directly fromethanolamine and, thus, appears to be a specific adaptation for glycine betaine accumulation during drought and salt-stress (Weretilnyke et al.,1995). The enzymes involved in the pathways illustrated are numbered: (1) glycerol-3-phosphate (G3P) is converted to phosphatidic acid bythe successive actions of G3P acyltransferase and 1-acylglycerol-3-phosphate acyltransferase; (2) phosphatidic acid phosphatase; (3) 1,2-diacylglycerol:choline phosphotransferase; (4) diacylglycerol acyltransferase; (5,6) carnitine palmitoyl transferases I and II; (7) trimethylamineoxygenase; (8) choline monoxygenase; (9) betaine aldehyde dehydrogenase; (10) P-choline phosphatase. TMA, trimethylamine; P-choline,phosphocholine.
greater relative capacity for betaine accumulation and their
via phosphatidylcholine hydrolysis has no known function
greater salt tolerance (McDonnell and Wyn Jones, 1988).
other than the synthesis of acetylcholine in neural tissue (Billah
Phosphatidylcholine hydrolysis is also responsible for
and Anthes, 1990). We propose that phosphatidylcholine
diacylglycerol production in plants that store large quantities
hydrolysis via the back reaction of CPT may provide an
of triacylglycerols in their seeds (Gur and Harwood, 1991).
endogenous source of choline for TMAO synthesis in animals.
Triacylglycerol sometimes constitutes as much as 80 % of the
Hydrolysis of dietary phosphatidylcholine by phospholipases
dry mass of the seeds. Triacylglycerols are also among the
C and D may also be an important source of free choline for
most common form of energy reserve in animals, but
TMAO synthesis (Wakelam et al., 1993). Hydrolysis of labeled
diacylglycerol ethers are also frequently stored. Many animals
phosphatidylcholine by phospholipase C in the gut of larvae of
store acylglycerols as metabolic fuel reserves that can, during
the dragonfly Aeshna cyanea resulted in recovery of labeled
starvation, migration, reproduction or egg development, be
products in various forms, including acylglycerols and glycine
mobilized and oxidized to drive metabolic processes.
Phosphatidylcholine hydrolysis, which is important during
Ordinarily, phospholipid synthesis takes precedence over
accumulation of acylglycerols, may serve as an endogenous
triacylglycerol synthesis in plants and animals when the
source of choline, resulting in TMAO accumulation in animals.
demand for accumulation of fuel stores is low. This ensures
The enzymes that are required for phosphatidylcholine
the maintenance of membrane turnover, an essential
hydrolysis are apparently well conserved, having been found
physiological process. However, during periods when storage
in mammals, molluscs and arthropods as well as the plants
of fuel is essential, such as preparation for seasonal reductions
mentioned above (Anfuso et al., 1995). Diacylglycerol and phosphatidylcholine
The predominant pathway for the biosynthesis of
triacylglycerol and diacylglycerol ether is the glycerol
phosphate pathway. Glycerol phosphate, a derivative ofglycolysis, is converted to diacylglycerol via phosphatidic acid
(Fig. 2). Diacylglycerol represents a branchpoint wherediacylglycerol can be either channeled into phospholipid
synthesis (i.e. phosphatidylcholine) or acylated to form
triacylglycerols. The final step towards the synthesis ofphosphatidylcholine is catalyzed by 1,2-diacylglycerol:choline
phosphotransferase (CPT). CPT activity governs thepartitioning of diacylglycerol into either phosphatidylcholineor acyglycerol pools (Jackowski et al., 2000). The backreaction of CPT can occur to a significant extent so that
diacylglycerol is formed from phosphatidylcholine, releasingfree choline (for a review, see Gur and Harwood, 1991).
phosphatidylcholine hydrolysis is an important second
Fig. 3. Trimethylamine oxide content (y, mmol kg–1; see Fig. 4) in
messenger (Billah and Anthes, 1990; Wakelam et al., 1993).
mantle muscle tissue is significantly correlated with digestive gland
In fact, during hypo-osmotic cell volume regulation,
lipid content (x) in cephalopods (y=2.90x1.18, r=0.59, P<0.05). We
swelling results in membrane turnover and
were unable to analyze trimethylamine oxide (TMAO) content as a
phosphatidylcholine hydrolysis, resulting in the formation of
function of total lipid content because of the variable presentation of
diacylglycerol. Diacylglycerol activates phosphokinase C
lipid data in the literature. Lipid data were taken from Blanchier and
which, in turn, stimulates the release of osmolytes in a range
Boucaud-Camou (1984), Hayashi (1989, 1996), Hayashi and
of organisms from sharks (Musch and Goldstein, 1990) to
Kawasaki (1985), Kristensen (1984), Phillips et al. (2001),
algae (Thompson, 1994). Cell volume regulation via
Piatkowski and Hagen (1994), Pollero and Iribarne (1988) and
phosphatidylcholine hydrolysis under hyperosmotic stress (i.e.
Semmens (1998). In many cases, lipid and TMAO data were taken
dehydration) could contribute to the release of free choline and
from different species, and possibly different maturity stages, of thesame genus. (1) Octopus; (2) Loligo; (3) Galiteuthis; (4) Sepia; (5)
subsequent betaine or TMAO accumulation in some animals
Illex; (6) Berryteuthis; (7) Moroteuthis; (8) Gonatopsis; (9)
(although some mechanism would be required to prevent
Todarodes; (10) Gonatus. There are conflicting reports regarding the
diacylglycerol from stimulating the release of osmolytes in this
digestive gland lipid content of Thysanoteuthis, a squid that also
case). For example, TMAO was shown to accumulate during
contains high TMAO concentrations (Hayashi, 1996; Yuneva et al.,
dehydration in frog gastronemius muscle (Wray and Wilkie,
1994). Many of the genera plotted here are closely related to each
1995), although the authors postulate that TMAO accumulated
other (see Fig. 4). Therefore, phylogenetic independence of the data
to counteract increased urea concentrations. Choline produced
Trimethylamine oxide in marine animals
in productivity, migrations or reproductive events,
Cod (gadiform teleosts) are sought commercially for their
diacylglycerol is preferentially channeled towards
abundant liver oil and have high concentrations of TMAO in
triacylglycerol synthesis or is converted to diacylglycerol
their muscle tissue (Gillett et al., 1997; Agustsson and Strøm,
ether. In at least one case, phosphatidylcholine itself is used as
1981). Among Antarctic fishes, Dissostichus sp. has the
a seasonal lipid reserve (Hagen et al., 1996). During such
highest concentrations of both body and liver lipids (Eastman,
periods, diacylglycerol production may be enhanced by
1988; Friedrich and Hagen, 1994) and TMAO (Raymond and
phosphatidylcholine hydrolysis via phospholipases or through
DeVries, 1998). Elasmobranchs (sharks) generally contain
the back reaction of CPT. The regulation of the enzymes
high levels of both liver lipids (Baldridge, 1970; Bone and
involved in these processes is poorly understood, but probably
Roberts, 1969) and TMAO (Withers et al., 1994). For example,
involves hormonal changes associated with ontogenetic,
Squalus sp. and Somniosus sp. have among the highest reported
TMAO levels (Anthoni et al., 1991a; Goldstein et al., 1967)and are sought commercially for the high concentrations ofdiacylglycerol ether (DAGE) in their livers (Hallgren and
Acylglycerol and TMAO: correlation
Stallberg, 1974; Kang et al., 1997). Somniosus microcephalus
A general correlation exists between the concentration of
has even been implicated in trimethylamine poisoning
TMAO (and betaine) in muscle tissue and lipid, particularly
(Anthoni et al., 1991a). Holocephalans, a generally deep-living
diacylglycerol ethers and triacylglycerols, levels in the bodies of
subclass of Chondrichthyes, also contain large quantities of
marine animals. TMAO and lipid concentration both appear to
methylamines (both betaine and TMAO) and lipid (Hayashi
be correlated with habitat depth, latitude, season, lifestyle (e.g.
and Takagi, 1980; Hebard et al., 1982; Bedford et al., 1998).
benthic versus pelagic) and ontogeny or size (for reviews, see
Among invertebrates, the correlation between TMAO and
Hebard et al., 1982; Sargent, 1976, 1989). Both TMAO and lipid
acylglycerol levels is most notable in the cephalopods (Fig. 3),
also appear conspicuously within the same phylogenetic groups.
especially in the deep-sea squid families Onychoteuthidae
Fig. 4. Mantle muscle trimethylamine oxide (TMAO, mmol kg–1) measured according to Wekell and Barnett (1991), minimum depth ofoccurrence (MDO, m) and capture depth (CD, m) of cephalopods listed according to their phylogenetic associations (see Carlini and Graves,1999). Values are corrected for the dilution of tissue with extracellular ammonium concentrations in some species (see text). Nodes arenumbered for reference in the text. Numbers in parentheses are from Hebard et al. (1982) or Kelly and Yancey (1999).
gonatid squids accumulates throughout their life and is thought
y = 75x0.33, r = 0.92
to fuel an extended egg-brooding period (Seibel et al., 2000). y = 2.5x0.25, r = 0.89
The high TMAO values reported for gonatid squids were found
y = 0.27x0.47, r = 0.74
in adult specimens. Preliminary measurements suggest that theconcentration of TMAO is much lower in smaller individuals
of the species (Fig. 5) and, as for lipids, that TMAO
Given the ontogenetic descent to great depths undertaken
by gonatid squids, an ontogenetic increase in TMAO
concentration is consistent with the hypothesis that TMAO
counteracts the effects of high hydrostatic pressure on proteins
(Kelly and Yancey, 1999). However, values for othercephalopods (Hebard et al., 1982) (Fig. 4) suggest that TMAO
content is not related to depth (although data on glycine betaineand other methylamines would be helpful to analyze
this hypothesis fully). For example, ommastrephids are
predominantly shallow-living squid that contain large amounts
of TMAO (100–335 mmol l–1; Fig. 4, node 13) (Hebard et al.,
1982). Ommastrephids accumulate lipid, up to 6 % of body
Fig. 5. Mantle muscle trimethylamine oxide (TMAO) content
mass, which is thought to fuel a reproductive horizontal
(mmol kg–1; squares) increases (b=0.33) in proportion to digestive
migration (Takahashi, 1960; Clarke et al., 1994). Incidentally,
gland mass (% body mass; circles; b=0.25) and lipid mass (% body
this family apparently has a ‘different’ smell from other squids
mass; plus signs; b=0.47) through ontogeny (size; g) in gonatid squid
and is known to cause allergic reactions in some people
(Cephalopoda). The data for TMAO content are from Kelly and
(Vecchione, 1994). Both phenomena are perhaps related to
Yancey (1999) and B. A. Seibel (unpublished results). Body masses
for the TMAO scaling analysis (squares) for Gonatopsis borealis and
Bathyteuthidae and Histioteuthidae, midwater squid families
Berryteuthis magister were estimated at the mean adult body mass
known to have large oily livers, were also found to accumulate
for each species (Hayashi, 1989; Hayashi and Yamamoto, 1987).
TMAO (Fig. 4). Furthermore, no relationship between TMAO
Digestive gland and lipid masses were taken from various sources:
content and minimum or capture depth was found for octopods
Gonatus onyx (B. A. Seibel, personal observation), Gonatus fabricii(Arkhipkin and Bjorke, 1999), Gonatopsis borealis (Hayashi, 1989)
over a wide depth range (Fig. 4, node 8). Incirrate octopods
and Berryteuthis magister (Hayashi and Yamamoto, 1987). All
(Fig. 4, node 11) have low TMAO levels at all depths. Cirrate
species appear to fall on the same scaling line despite differences in
octopods (Fig. 4, node 16) also have fairly low levels of
maximum body sizes. The apparent correlation between lipid content
TMAO, but slightly higher levels of betaine (>50 mmol kg–1)
(or digestive gland mass) and TMAO content in gonatid squid is
(Yin and Yancey, 2000). The few shallow-living octopods
hypothesized to result from the requirement for phosphatidyl choline
measured have had very low lipid contents (O’Dor and
hydrolysis to produce diacylglycerol for lipid storage. The choline
Webber, 1986; Pollero and Iribarne, 1988). Lipid levels have
not been measured in cirrate octopods. The general increase inTMAO concentration with depth observed in a variety of
(Moroteuthis robusta) (Fig. 4; node 7) and Gonatidae (Fig. 5)
animals (Kelly and Yancey, 1999) may be related to the
(Fig. 4, node 10). While most cephalopods studied to date have
tendency for deep-living species to accumulate lipids (e.g.
very low lipid concentrations (O’Dor and Webber, 1986) and
little ability to metabolize lipids (Hochachka, 1994), gonatid
and onychoteuthid squids have acylglycerol contents as high
(approximately 300 mmol kg–1) in their tissues for buoyancy.
as 25 % of their body mass (Hayashi and Kawasaki, 1985;
Although previous methods measuring ammonium did not
Hayashi et al., 1990; Phillips et al., 2001). These two families
distinguish between ammonium and methylamines (Sanders
also contain the highest TMAO concentrations among
and Childress, 1988), our recent analysis using high-
cephalopods (Fig. 4) (Hebard et al., 1982; Kelly and Yancey,
performance liquid chromatography confirmed the existence of
1999). Some gonatid squids apparently contain high
high ammonium concentrations in most midwater squid
concentrations of glycine betaine as well (Shirai et al., 1997).
species (B. A. Seibel, unpublished data). Such species appear
Both gonatid and onychoteuthid squid are known to
to have special vacuolated tissue in which ammonium is
undertake ontogenetic vertical migrations whereby successive
sequestered, presumably out of contact with intracellular
developmental stages occupy progressively greater depths.
macromolecules (Voight et al., 1994). If this sequestration is
This migration is believed to end at depths greater than 1500 m,
incomplete, however, TMAO could be required to counteract
at which spawning and, at least in some cases, egg-brooding
the toxic effects of ammonium on enzymes in ammoniacal
take place (Jackson and Mladenov, 1994; Arkhipkin and
squid. Preliminary measurements suggest that ammonium does
Bjorke, 1999; Seibel et al., 2000). The high lipid content in
depress the activity of octopine dehydrogenase from
Trimethylamine oxide in marine animalsHistioteuthis heteropsis muscle tissue. However, TMAO did
Time course of methylamine accumulation
not effectively counteract this depression (B. A. Seibel,
Although some groups, such as elasmobranchs, have
unpublished data). Furthermore, most species with high
specific adaptations for retention of TMAO to counteract urea
TMAO concentrations are negatively buoyant and do not
toxicity, other groups may simply allow TMAO to accumulate
accumulate ammonium. In the case of gonatids, the
while it is available and thus reap the benefits of its
ontogenetic stages with high TMAO values also have lipid
compatibility and protein-stabilizing attributes. The time
contents sufficient to provide neutral buoyancy and do not
course of glycine betaine accumulation in wheat plants during,
require ammonium or TMAO to provide lift.
and subsequent to, periods of leaf expansion is consistent with
It should be pointed out that cirrate and some pelagic
this explanation (Hitz et al., 1981; McDonnell and Wyn Jones,
incirrate octopods have extensive extracellular gelatinous
1988). The most rapid betaine accumulation in the leaves of
tissue. We made every effort to remove the external gelatinous
unstressed wheat plants, and initially in those of salt-stressed
tissue, but some dilution of the TMAO content is expected in
wheat plants, is at the time of greatest leaf expansion and,
these species. Inclusion of the extracellular ammonium
consequently, membrane development and membrane lipid
vacuoles in the analyzed tissue of some midwater squid
(e.g. phosphatidylcholine) turnover. When growth (glycolypid
certainly diluted the tissue homogenates as well. Assuming that
biosynthesis) plateaus, betaine concentrations slowly decline
all ammonium in such species is contained as a 500 mmol l–1
solution in extracellular vacuoles (Voight et al., 1994) and that
The relatively low TMAO concentrations (48 mmol kg–1)
all TMAO measured is intracellular, we have corrected these
that we recently measured in a gonatid squid that was
values (Fig. 4) using measured ammonium and protein
brooding an egg mass are also consistent with this model (B.
contents (B. A. Seibel, unpublished data). Bathyteuthis berryi
A. Seibel, unpublished data). This squid was using, rather
does not possess ammonium as previously reported, but does
than storing, lipid and had lost the TMAO that it had
contain high concentrations of an as yet unidentified
presumably accumulated prior to spawning (Fig. 5).
nitrogenous cation, possibly TMA (B. A. Seibel, unpublished
Dehydrated frog muscle initially accumulated methylamines
data) (Voight et al., 1994). We believe that this cation is
in concert with rising urea concentrations (Wray and Wilkie,
intracellular and so no correction to the TMAO value in Fig. 4
1995). However, urea concentrations continued to rise with
has been applied to this species. Galiteuthis phyllura
continued dehydration, while methylamine levels reached a
accumulates ammonium in a specialized coelomic chamber
well out of contact with muscle tissue. No correction wasapplied to this species either. TMAO was not found in thecoelomic fluid of Galiteuthis phyllura. Adaptive significance of methylamines
We recently measured high TMAO concentrations in
The role of phosphatidylcholine hydrolysis in the
Clione antarctica (112 mmol kg–1) (B. A. Seibel, unpublished
production, as well as the time course of accumulation, of
data), a shallow-living Antarctic pteropod mollusc known to
betaine led Hitz et al. (1981) to question the adaptive
store high concentrations of DAGE within its body (Kattner
significance of betaine accumulation in wheat and barley.
et al., 1998; Phleger et al., 1997). Clione antarctica
Betaine accumulation may only be a side-effect of accelerated
feeds exclusively on the thecosomatous pteropod Limacina
turnover of phospholipid head groups during stress and not a
helicina, which accumulates large quantities of
specific adaptation to stress from which some benefit accrues
dimethylsulfoniopropionate (DMSP) in its body (Levasseur et
to the stressed leaf (Hitz et al., 1981). In oysters, betaine
al., 1994) directly from phytoplankton in its diet. DMSP is the
synthesis and accumulation may be a consequence of cell
sulphidic analog of the nitrogenous glycine betaine and
membrane restructuring during cell volume changes resulting
may be the precursor of the sulfonium analogue of
in phosphatidylcholine hydrolysis (cf. Dragolovich, 1994;
phosphatidylcholine in some phytoplankton (Kates and
Musch and Goldstein, 1990). Similarly the temporary
Volcani, 1996). Like trimethylamine oxide, DMSP is known
accumulation of trimethylamines in frog muscle during
to confer some protection against osmotic and temperature
dehydration may also reflect membrane turnover during
stress in phytoplankton (Nishiguchi and Somero, 1992).
hyperosmotic cell volume regulation (see Wray and Wilkie,
Fishes that feed on Limacina helicina are often inflicted
1995) and not necessarily a response to increased urea
with ‘blackberry feed’, a foul-smelling and aesthetically
concentrations. The adaptive significance of DMSP
displeasing condition resulting from the breakdown of DMSP
accumulation in algal cells has also been called into question
to dimethylsulfide, subsequently accumulated in the fish’s
by Stefels (2000), who noted that changes in the concentration
tissues (Levasseur et al., 1994). Although the TMAO
of DMSP in algal cells upon salt-stress are the result of
measured in Clione antarctica may be related to the large lipid
metabolic changes rather than active regulatory mechanisms.
stores, some connection to the DMSP concentrations in
He suggested that DMSP may be considered as a compatible
Limacina helicina cannot be ruled out. The synthetic pathways
solute, but that it is not osmoticum in the strict sense of being
of DMSP and betaine are linked (Mulholland and Otte, 2000).
responsible for osmotic balance. Perhaps TMAO accumulation
Trimethylamine oxide levels in Limacina helicina have not
in marine animals reflects, in part, the requirements for
Fatty liver in sharks
organisms. TMAO may simply be accumulated as a
In mammals, dietary choline deficiency prevents
compatible solute in quantities reflecting the amount of lipid
phosphatidylcholine synthesis and may leave excess
stored in the body. Conversely, among sharks, the requirement
diacylglycerol, produced in the glycerol phosphate pathway, to
for TMAO accumulation may deplete available choline levels,
be channeled towards triacylglycerol for storage in the liver
thus limiting the production of phosphatidylcholine and
(Fig. 2). This condition, known as ‘fatty liver’, may be a
shunting excess diacylglycerol, produced in the glycerol
chronic condition in sharks. The requirements for TMAO
phosphate pathway, towards storage in the liver. Sharks alone
accumulation to counteract urea toxicity may limit the
among marine animals may possess specific adaptations for
availability of choline for phosphatidylcholine synthesis. As a
retention, and possibly for production via centralization of
consequence, lipid may accumulate in the liver. The loss of
fatty-acid oxidation, of TMAO. We do not rule out the
extra-hepatic fatty acid oxidation may further contribute to
possibility that TMAO is strongly selected for its protein-
fatty liver in sharks. Freshwater stingrays alone among
stabilizing attributes in some other animal groups, possibly
elasmobranchs possess the ability to oxidize fatty acids extra-
resulting in lipid accumulation as a metabolic byproduct. It is
hepatically. Because they have no need to accumulate urea as
also possible that that diacylglycerol and TMAO levels are
an osmolyte, freshwater stingrays also do not accumulate
linked, as we have proposed, but that the maintenance of high
TMAO in their tissues. This led Ballantyne and Moon (1986)
TMAO levels reflects retention adaptations in response to some
to postulate a relationship between extra-hepatic β-oxidation
cellular perturbant. However, no obvious protein-destabilizing
and the absence of urea and TMAO accumulation. One
agent has been identified in the cephalopods examined here
possible cause is competition for carnitine which, like choline,
that would warrant the observed accumulation of TMAO.
may be oxidized to TMAO (Marzo and Curti, 1997).
Because of the paucity of data on methylamines other than
β-oxidation may allow all available carnitine to be
TMAO, we hesitate to rule out the possibility that hydrostatic
converted to TMAO. By limiting β-oxidation in non-hepatic
pressure selects for high methylamine concentrations in deep-
tissue, sharks may also decrease the competition for
sea organisms. We also cannot rule out the possibility that
diacylglycerol for lipid accumulation in the liver. Limited
pathways for the accumulation of lipid and TMAO are not
extra-hepatic fatty acid oxidation capacity in squid (Ballantyne
coupled in some cases. However, we feel that the link between
et al., 1981) may similarly preadapt them for seasonal and
phosphatidylcholine hydrolysis and trimethylamine oxide
ontogenetic acylglycerol accumulation. However, the only
accumulation should be considered a competing hypothesis
genus for which the capacity for fatty acid oxidation has been
measured (Loligo) does not appear to accumulate lipid orTMAO. The lipid-rich livers of sharks are generally attributed
We thank the Monterey Bay Aquarium for allowing us to
a buoyancy role (Malins and Barone, 1970; Phleger, 1998;
participate in collection cruises. We thank G. N. Somero and
Wetherbee and Nichols, 2000). Although the low-density oils,
two anonymous reviewers for comments and constructive
especially DAGE and squalene, do provide lift, they may
criticism. This research was funded in part by the Rosenstiel
simply be a beneficial end-product of TMAO production in
School of Marine and Atmospheric Science Postdoctoral
Fellowship and National Institute of Environmental Health
Although this ‘fatty liver’ scenario could, in theory, also
Sciences Marine and Freshwater Biomedical Science Center
apply to animal groups other than elasmobranchs, our inability
to identify a cellular perturbant that consistently explains thedistribution of TMAO in cephalopods and other marineanimals causes us to reject this possibility. Among marine
References
animals, methylamine accumulation as an adaptation for
Agustsson, I. and Strøm, A. R. (1981). Biosynthesis and turnover of
trimethylamine oxide in the teleost cod, Gadus morhua. J. Biol. Chem. 256,
macromolecular stabilization has, in our opinion, been
convincingly demonstrated only in sharks. Anfuso, C. D., Sipione, S., Lupo, G. and Alberghina, M. (1995).
Fatty liver in mammals often results in cancerous tumor
Evolutionary comparison of enzyme activities of phosphatidylcholinemetabolism in the nervous system of an invertebrate (Loligo pealei), lower
production (Goshal and Farber, 1984; Locker et al., 1986). It
vertebrate (Mustelus canis) and the rat. Comp. Biochem. Physiol. 112B,
may be no coincidence that shark livers (and squid digestive
glands for that matter) also contain large quantities of
Anthoni, U., Christophersen, C., Gram, L., Nielsen, N. H. and Nielsen, P.
(1991a). Poisonings from flesh of the greenland shark, Somniosus
squalamine, derived from squaline lipids and/or diacylglycerol
microcephalus, may be due to trimethylamine. Toxicon 29, 1205–1212.
ethers. Both squalamine and diacylglycerol ethers have
Anthoni, U., Christophersen, C., Hougaard, L. and Nielsen, P. H. (1991b).
recently been shown to inhibit tumor development by limiting
Quaternary ammonium compounds in the biosphere – an example of a versatile adaptive strategy. Comp. Biochem. Physiol. 99B, 1–18.
vascular growth (Sills et al., 1998). Arkhipkin, A. I. and Bjorke, H. (1999). Ontogenetic changes in
morphometric and reproductive indices of the squid Gonatus fabricii (Oegopsida, Gonatidae) in the Norwegian Sea. Polar Biol. 22, 357–365. Concluding remarks Baker, J. R., Streuempler, A. and Chaykin, S. (1963). A comparative study
of trimethylamine-N-oxide biosynthesis. Biochim. Biophys. Acta 71, 58–64.
We suggest that hydrolysis of phosphatidylcholine
Baldridge, D. H. J. (1970). Sinking factors and average densities of Florida
contributes to TMAO accumulation in many marine
sharks as functions of liver buoyancy. Copeia 4, 744–754. Trimethylamine oxide in marine animalsBallantyne, J. S. (1997). Jaws: The inside story. The metabolism of Hayashi, K. (1996). Composition and distribution of lipids in different tissues
elasmobranch fishes. Comp. Biochem. Physiol. 118B, 703–742.
of the arrow squid, Loligo bleekeri. Fish. Sci. 62, 84–87. Ballantyne, J. S., Hochachka, P. W. and Mommsen, T. P. (1981). Studies Hayashi, K. and Kawasaki, K. (1985). Unusual occurrence of diacyle
on the metabolism of the migratory squid, Loligo opalescens: enzymes of
glyceryl ethers in liver lipids from two species of Gonatid squids. Bull. Jap.
tissues and heart mitochondria. Mar. Biol. Lett. 2, 75–85. Soc. Scient. Fish. 51, 593–597. Ballantyne, J. S. and Moon, T. W. (1986). The effects of urea, Hayashi, K., Kishimura, H. and Sakurai, Y. (1990). Level and composition
trimethylamine oxide and ionic strength on the oxidation of acyl carnitines
of diacyl glyceryl ethers in the different tissues and stomach contents of
by mitochondria isolated from the liver of the little skate Raja erinacea. J.
giant squid Moroteuthis robusta. Nippon Suisan Gakkaishi 56, 1635–1639. Comp. Physiol. 156, 845–851. Hayashi, K. and Takagi, T. (1980). Composition of diacyl glycerol ethers in Bedford, J. J., Harper, J. L., Leader, J. P., Yancey, P. H. and Smith, R.
the liver lipids of ratfish, Hydrolagus novaezealandiae. Bull. Jap. Soc.A. J. (1998). Betaine is the principal counteracting osmolyte in tissue of the Scient. Fish. 46, 855–861.
elephant fish, Callorhincus millii (Elasmobranchii, Holocephali). Comp.Hayashi, K. and Yamamoto (1987). Distribution of diacyl glycerol ethers in Biochem. Physiol. 119B, 521–526.
the different tissues and stomach contents of gonatid squid BerryteuthisBillah, M. M. and Anthes, J. C. (1990). The regulation and cellular functions magister. Nippon Suisan Gakkaishi 53, 137–140.
of phosphatidylcholine hydrolysis. Biochem. J. 269, 281–291. Hebard, C. E., Flick, G. J. and Martin, R. E. (1982). Occurrence and Blanchier, B. and Boucaud-Camou, E. (1984). Lipids in the digestive gland
significance of trimethylamine oxide and its derivatives in fish and shellfish.
and the gonad of immature and mature Sepia officinalis (Mollusca:
In Chemistry and Biochemistry of Marine Food Products (ed. R. E. Martin,
Cephalopoda). Mar. Biol. 80, 39–43.
G. J. Flick, C. E. Hebard and D. R. Ward), pp. 149–304. Westport, CT: AVI
Bone, Q. and Roberts, B. L. (1969). The density of elasmobranchs. J. Mar. Biol. Ass. U.K. 49, 409–425. Hitz, W. D., Rhodes, D. and Hanson, A. D. (1981). Radiotracer evidence Carlini, D. B. and Graves, J. E. (1999). Phylogenetic analysis of cytochrome
implicating phosphoryl and phosphatidyl bases as intermediates in betaine
C oxidase I sequences to determine higher-level relationships within the
synthesis by water-stressed barley leaves. Plant Physiol. 68, 814–822.
coleoid cephalopods. Bull. Mar. Sci. 64, 57–76. Hochachka, P. W. (1994). Oxygen efficient design of cephalopod muscle Childress, J. J. and Nygaard, M. H. (1974). The chemical composition and
metabolism. In Physiology of Cephalopod Molluscs: Lifestyle and
relative buoyancy of midwater crustaceans as a function of depth off
Performance Adaptations (ed. R. K. O’Dor, H. O. Pörtner and D. L.
Southern California. Mar. Biol. 27, 225–238.
MacMillan), pp. 61–67. Basel, Switzerland: Gordon & Breach Publishers. Clarke, A., Rodhouse, P. G. and Gore, D. J. (1994). Biochemical Hunt, J. C. and Seibel, B. A. (2000). Life history of Gonatus onyx
composition in relation to the energetics of growth and sexual maturation
(Teuthoidea: Cephalopoda): ontogenetic changes in habitat, behavior and
in the ommastrephid squid Illex argentinus. Phil. Trans. R. Soc. Lond. B
physiology. Mar. Biol. 136, 543–552. 344, 201–212. Jackowski, S., Wang, J. and Baburina, I. Cohen, J. J., Krupp, M. A. and Chidsey, C. A. (1958). Renal conservation
phosphatidylcholine biosynthetic pathway modulates the distribution of
of trimethylamine oxide by the spiny dogfish, Squalus acanthias. Am. J.
fatty acids into glycerolipids in prolifering cells. Biochim. Biophys. ActaPhysiol. 42, 319–327. 1483, 301–315. Dolphin, C. T., Janmohamed, A., Smith, R. L., Shephard, E. A. and Jackson, G. D. and Mladenov, P. V. (1994). Terminal spawning in the Phillips, I. R. (1997). Missense mutation in flavin-containing mono-
deepwater squid Moroteuthis ingens (Cephalopoda: Onychoteuthidae). J.
oxygenase 3 gene, FMO3, underlies fish-odour syndrome. Nature GeneticsZool., Lond. 234, 189–201. Kang, S. J., Lall, S. P. and Ackman, R. G. (1997). Digestion of the 1-O- Dragolovich, J. (1994). Dealing with salt stress in animal cells: the role
alkyl diacylglycerol ethers of Atlantic dogfish liver oils by Atlantic salmon,
and regulation of glycine betaine concentrations. J. Exp. Zool. 268, Salmo salar. Lipids 32, 19–30. Kates, M. and Volcani, B. E. (1996). Biosynthetic pathways for Eastman, J. T. (1988). Lipid storage systems and the biology of two neutrally
phosphatidylsulfocholine, the sulfonium analogue of phosphatidylcholine,
buoyant Antarctic notothenioid fishes. Comp. Biochem. Physiol. 90B,
in diatoms. In Biological and Environmental Chemistry of DMSP andRelated Sulfonium Compounds (ed. R. P. Kien, P. T. Visscher, M. D. Keller
Friedrich, C. and Hagen, W. (1994). Lipid contents of five species of
and G. O. Kirst), pp. 109–119. New York: Plenum Press.
notothenioid fish from high-Antarctic waters and ecological implications. Kattner, G., Hagen, W., Graeve, M. and Albers, C. (1998). Exceptional Polar Biol. 14, 359–369.
lipids and fatty acids in the pteropod Clione limacina (Gastropoda) from
Gillett, M. B., Suko, J. R., Santoso, F. O. and Yancey, P. H. (1997).
both polar oceans. Mar. Chem. 61, 219–228.
Elevated levels of trimethylamine oxide in muscles of deep-sea gadiform
Kelly, R. H. and Yancey, P. H. (1999). High contents of trimethylamine oxide
teleosts: a high-pressure adaptation? J. Exp. Zool. 279, 386–391.
correlating with depth in deep-sea teleost fishes, skates and decapod
Goldstein, L. and Funkhouser, D. (1972). Biosynthesis of trimethylamine
crustaceans. Biol. Bull. 196, 18–25.
oxide in nurse shark, Ginglymostoma cirratum. Comp. Biochem. Physiol.Kloiber, O., Banjac, B. and Drewes, L. R. (1988). Protection against acute 42A, 51–57.
hyperammonemia: the role of quaternary amines. Toxicology 49, 83–90. Goldstein, L., Hartman, S. C. and Forster, R. P. (1967). On the origin of Krall, J. P., Edwards, G. E. and Andreo, C. S. (1989). Protection of pyruvate
trimethylamine oxide in the spiny dogfish, Squalus acanthias. Comp.
kinase from maize against cold lability by compatible solutes. Plant Physiol.Biochem. Physiol. 21, 719–722. 89, 280–285. Goldstein, L. and Palatt, P. J. (1974). Trimethylamine oxide excretion rates Kristensen, T. K. (1984). Biology of the squid Gonatus fabricii (Lichtenstein,
in elasmobranchs. Am. J. Physiol. 227, 1268–1271.
1818) fom west Greeland waters. Meddelelser om Gronland, Bioscience 13, Goshal, A. K. and Farber, E. (1984). The induction of liver cancer by dietary
deficiency of choline and methionine without added carcinogens. Levasseur, M., Keller, M. D., Bonneau, E., D’Amours, D. and Bellows, W. Carcinogenesis 5, 1367–1370. K. (1994). Oceanographic basis of a DMS-related Atlantic Cod (Gadus Gur, M. I. and Harwood, J. L. (1991). Lipid Biochemistry: An Introduction. morhua) fishery problem: blackberry feed. Can. J. Fish. Aquat. Sci. 51, Hagen, W., Van Vleet, E. S. and Kattner, G. (1996). Seasonal lipid storage Locker, J., Reddy, T. V. and Lombardi, B. (1986). DNA methylation and
as overwintering strategy of Antarctic krill. Mar. Ecol. Prog. Ser. 134,
hepatocarcinogenesis in rats fed a choline-devoid diet. Carcinogenesis 7, Hallgren, B. and Stallberg, G. (1974). 1-O-(2-Hydroxyalkyl) glycerols Malins, D. C. and Barone, A. (1970). Glyceryl ether metabolism: regulation
isolated from Greenland shark liver oil. Acta Chem. Scand. 28B,
of buoyancy in dogfish Squalus acanthias. Science 167, 79–80. Marzo, A. and Curti, S. (1997). L-Carnitine moiety assay: an up-to-date Hanson, A. D. and Rhodes, D. (1983). 14C tracer evidence for synthesis of
reappraisal covering the commonest methods for various applications. J.
choline and betaine via phosphoryl base intermediates in salinized sugarbeet
Chromatogr. 702, 1–20.
leaves. Plant Physiol. 71, 692–700. McDonnell, E. and Wyn Jones, R. G. (1988). Glycinebetaine biosynthesis Hayashi, K. (1989). Occurrence of diacyl glyceryl ethers in liver lipids of
and accumulation in unstressed and salt-stressed wheat. J. Exp. Bot. 39,
Gonatid squid Gonatopsis borealis. Nippon Suisan Gakkaishi 55, Minana, M., Hermenegildo, C., Llansola, M., Montoliu, C., Grisolia, S. and Felip, V. (1996). Carnitine and choline derivatives containing a Seafood Enzymes (ed. N. F. Haard and B. K. Simpson), pp. 167–190. New
trimethylamine group prevent ammonia toxicity in mice and glutamate
toxicity in primary cultures of neurons. J. Pharmacol. Exp. Ther. 279, Stefels, J. (2000). Physiological aspects of the production and conversion of
DMSP in marine algae and higher plants. J. Sea Res. 43, 183–197. Mulholland, M. M. and Otte, M. L. (2000). Effects of varying sulphate and Strom, A. R. (1979). Biosynthesis of trimethylamine oxide in calanoid
nitrogen supply on DMSP and glycine betaine levels in Spartina anglica. J.
copepods. Seasonal changes in trimethylamine monooxygenase activity. Sea Res. 43, 199–207. Mar. Biol. 51, 33–40. Musch, M. W. and Goldstein, L. (1990). Hypotonicity stimulates Summers, P. S. and Weretilnyk, E. A. (1993). Choline synthesis in spinach
phosphatidylcholine hydrolysis and generates diacylglycerol in
in relation to salt stress. Plant Physiol. 103, 1269–1276.
erythrocytes. J. Biol. Chem. 265, 13055–13059. Takahashi, T. (1960). Studies on the utilization of the cuttlefish, Nishiguchi, M. K. and Somero, G. N. (1992). Temperature- and Ommastrephes sloani pacificus. III. The seasonal variations in the
concentration dependence of compatibility of the organic osmolyte β-
gravimetric constitution and chemical composition of the various parts of
dimethylsulfoniopropionate. Cryobiology 29, 118–124.
the body. Bull. Jap. Soc. Scient. Fish. 26, 95–98. Nuccio, M. L., Russell, B. L., Rathinasabapathi, B., Gage, D. A. and Thompson, G. A. (1994). Mechanisms of osmoregulation in the green alga Hanson, A. D. (1998). The endogenous choline supply limits glycine Dunaliella salina. J. Exp. Zool. 268, 127–132.
betaine synthesis in transgenic tobacco expressing choline monooxygenase. Treacy, E. P., Akerman, B. R., Chow, L. M. L., Youil, R., Bibeau, C., Lin, Plant J. 16, 487–496. J., Bruce, A. G., Knight, M., Danks, D. M., Cashman, J. R. and Forrest, O’Dor, R. K. and Webber, D. M. (1986). The constraints on cephalopods: S. M. (1998). Mutations of the flavin-containing monooxygenase gene
why squid aren’t fish. Can. J. Zool. 64, 1591–1605.
(FMO3) cause trimethylaminuria, a defect in detoxication. Human Mol.Phillips, K. L., Jackson, G. D. and Nichols, P. D. (2001). Predation on Genet. 7, 839–845.
myctophids by the squid Moroteuthis ingens around Macquarie and Heard
Vecchione, M. (1994). Systematics and the lifestyle and performance of
Islands: stomach contents and fatty acid analyses. Mar. Ecol. Prog. Ser. 215,
cephalopods. In Physiology of Cephalopod Molluscs: Lifestyle andPerformance Adaptations (ed. H. O. Pörtner, R. K. O’Dor and D. L. Phleger, C. F. (1998). Buoyancy in marine fishes: direct and indirect role of
Macmillan,), pp. 179–191. Basel, Switzerland: Gordon & Breach
lipids. Am. Zool. 38, 321–330. Phleger, C. F., Nichols, P. D. and Virtue, P. (1997). Lipids and buoyancy in Voight, J. R., Pörtner, H. O. and O’Dor, R. K. (1994). A review of
Southern Ocean pteropods. Lipids 32, 1093–1100.
ammonia-mediated buoyancy in squids (Cephalopoda: Teuthoidea). Mar.Piatkowski, U. and Hagen, W. (1994). Distribution and lipid composition of Fresh. Behav. Physiol. 25, 193–203.
early life stages of the chranchiid squid Galiteuthis glacialis (Chun) in the
Wakelam, M. J. O., Briscoe, C. P., Stewart, A., Pettitt, T. R., Cross, M.
Weddell Sea, Antarctica. Antarctic Sci. 6, 235–239. J., Paul, A., Yule, J. M., Gardner, S. D. and Hodgkin, M. (1993). Pierce, S. K., Dragolovich, J. and Crombie, B. N. (1997). Variations in
Phosphatidylcholine hydrolysis: a source of multiple lipid messenger
intracellular choline levels may account for differences in glycine betaine
molecules. Biochem. Soc. Trans. 21, 374–378.
synthesis between conspecific oyster populations responding to
Wang, A. and Bolen, D. W. (1997). A naturally occurring protective system
hyperosmotic stress. J. Exp. Zool. 278, 283–289.
in urea-rich cells: mechanism of osmolyte protection of proteins against urea
Pollero, R. and Iribarne, O. (1988). Biochemical changes during the
denaturation. Biochemistry 30, 9101–9108.
reproductive cycle of the small Patagonian octopus, Octopus tehuelchus,
Weiher, B. and Komnick, H. (1997). Digestion of phosphatidylcholines,
D’Orb. Comp. Biochem. Physiol. 90B, 317–320.
absorption and esterification of lipolytic products by Aeshna cyaneaRaymond, J. A. (1994). Seasonal variations of trimethylamine oxide and urea
larvae as studied in vivo and in vitro. Arch. Insect Biochem. Physiol. 36,
in the blood of a cold-adapted marine teleost, the rainbow smelt. FishPhysiol. Biochem. 13, 13–22. Wekell, J. C. and Barnett, H. (1991). New method for analysis of Raymond, J. A. and DeVries, A. L. (1998). Elevated concentrations and
trimethylamine oxide using ferrous sulfate and EDTA. J. Food Sci. 56,
synthetic pathways of trimethylamine oxide and urea in some teleost fishes
of McMurdo Sound, Antarctica. Fish Physiol. Biochem 18, 387–398. Weretilnyk, E. A., Smith, D. D., Wilch, G. A. and Summers, P. S. (1995). Sanders, N. K. and Childress, J. J. (1988). Ion replacement as a buoyancy
Enzymes of choline synthesis in spinach: response of phospho-base N-
mechanism in a pelagic deep-sea crustacean. J. Exp. Biol. 138, 333–348.
methyltransferase activities to light and salinity. Plant Physiol. 109, Sargent, J. R. (1976). The structure, metabolism and function of lipids in
marine organisms. In Biochemical and Biophysical Perspectives in MarineWetherbee, B. M. and Nichols, P. D. (2000). Lipid composition of the liver Biology, vol. 3 (ed. D. C. Malins and J. R. Sargent), pp. 149–212. London:
oil of deep-sea sharks from the Chatham Rise, New Zealand. Comp.Biochem. Physiol. 125B, 511–521. Sargent, J. R. (1989). Ether-linked glycerides in marine animals. In Marine Withers, P. C., Morrison, G., Hefter, G. T. and Pang, T. (1994). Role of Biogenic Lipids, Fats and Oils (ed. R. G. Ackman), pp. 176–193. Boca
urea and methylamines in buoyancy of elasmobranchs. J. Exp. Biol. 188, Schlenk, D. and Li-Schlenk, R. (1994). Characterization of liver flavin- Wray, S. and Wilkie, D. R. (1995). The relationship between plasma urea
containing monooxygenase of the dogfish shark (Squalus acanthias) and
levels and some muscle trimethylamine levels in Xenopus laevis: A 31P and
partial purification of liver flavin-containing monooxygenase of the silky
14N nuclear magnetic resonance study. J. Exp. Biol. 198, 373–378.
shark (Carcharhinus falciformis). Comp. Biochem. Physiol. 109B, 655–664. Yancey, P. H., Clarke, M. E., Hand, S. C., Bowlus, R. D. and Somero, G. Seibel, B. A., Hochberg, F. G. and Carlini, D. B. (2000). Life history of N. (1982). Living with water stress: Evolution of osmolyte systems. Science Gonatus onyx: deep-sea spawning and post-spawning egg care. Mar. Biol.217, 1214–1222. 137, 519–526. Yancey, P. H., Fyfe-Johnson, A. L., Kelly, R. H., Walker, V. P. and Aunon, Semmens, J. M. (1998). An examination of the role of the digestive gland of M. T. (2001). Trimethylamine oxide counteracts effects of hydrostatic
two loliginid squids, with respect to lipid: storage or excretion? Proc. R.
pressure on proteins of deep-sea teleosts. J. Exp. Zool. 289, 172–176. Soc. Lond. B 265, 1685–1690. Yancey, P. H. and Siebenaller, J. F. (1999). Trimethylamine oxide stabilizes Shirai, T., Kikuchi, N., Matsuo, S., Uchida, S., Inada, H., Suzuki, T. and
teleost and mammalian lactate dehydrogeanses against inactivation by
Hirano, T. (1997). Taste components of boreo Pacific gonate squid
hydrostatic pressure and trypsinolysis. J. Exp. Biol. 202, 3597–3603. Gonatopsis borealis. Fish. Sci. 63, 772–778. Yancey, P. H. and Somero, G. N. (1980). Methylamine osmoregulatory Sills, A. K. J., Williams, J. I., Tyler, B. M., Epstein, D. S., Sipos, E. P.,
solutes of elasmobranch fishes counteract urea inhibition of enzymes. J. Exp.Davis, J. D., McLane, M. P., Pitchford, S., Cheshire, K., Gannon, F. H. Zool. 212, 205–213. and others, (1998). Squalamine inhibits angiogenesis and solid tumor Yin, M. and Yancey, P. H. (2000). Methylamine osmolytes dominate in deep-
growth in vivo and perturbs embryonic vasculature. Cancer Res. 58,
sea polychaetes, pycnogonids and octopods (abstract only). Am. Zool. 40, Somero, G. N. (1986). From dogfish to dogs: Trimethylamines protect Yuneva, T. V., Shchepkina, A. M. and Shul’man, G. E. (1994). Specificity
proteins from urea. News Physiol. Sci. 1, 9–12.
of lipids contain in calmar tissues from tropic Atlantic. GidrobiologicheskiiSotelo, C. G. and Rehbein, H. (2000). TMAO-degrading enzymes. In Zhurnal 30, 78–86.
- INFORMAÇÕES TÉCNICAS AOS PROFISSIONAIS DA SAÚDE Características Farmacológicas: Polizol é um anti-hemíntico polivalente, dotado de ação vermicida, ovicida e larvicida, exercendo sua atividade anti-helmíntica por bloqueio dos processos enzimáticos responsáveis pela absorção da glicose, com isto o nível de energia se torna inadequado à sobrevivência. Inicialmente os helmint
GUIDELINES FOR ELIGIBLE REIMBURSEMENTS GENERAL PLEASE COPY ALL ENCLOSURES FOR YOUR PERSONAL REFERENCE AND INCOME TAX RECORDS If you have not submitted the medical/dental expense to your insurance plan(s), please do so prior to submission on this Flexible Spending Account Reimbursement Request Form. If you apply for reimbursement of an expense that the IRS later determines to be inel
 greater relative capacity for betaine accumulation and their
via phosphatidylcholine hydrolysis has no known function
greater salt tolerance (McDonnell and Wyn Jones, 1988).
greater relative capacity for betaine accumulation and their
via phosphatidylcholine hydrolysis has no known function
greater salt tolerance (McDonnell and Wyn Jones, 1988).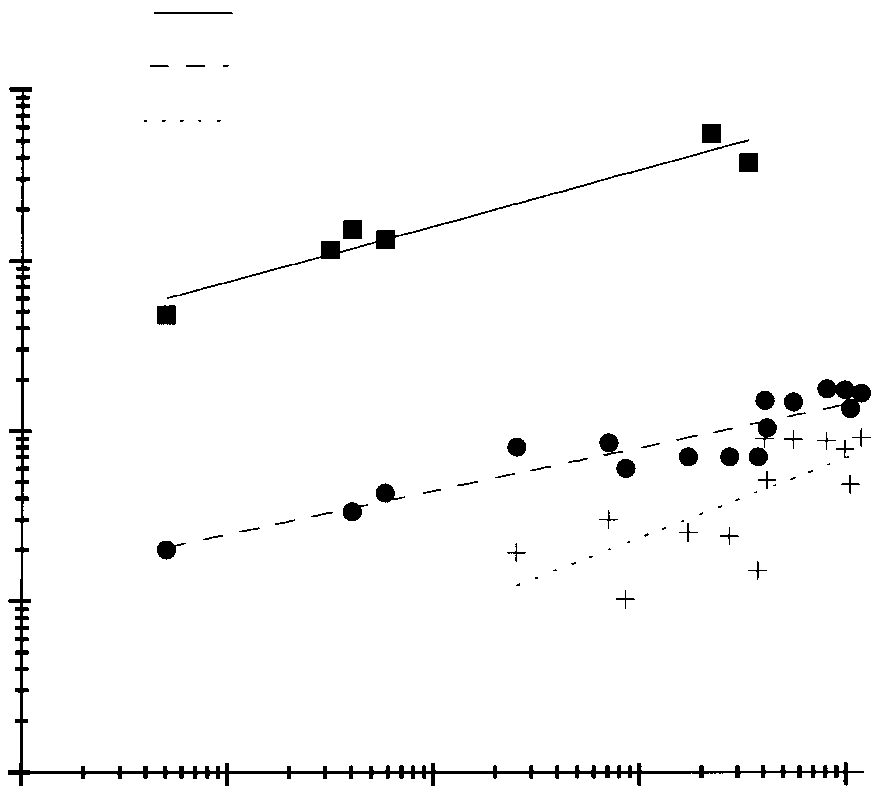 gonatid squids accumulates throughout their life and is thought
y = 75x0.33, r = 0.92
to fuel an extended egg-brooding period (Seibel et al., 2000).
gonatid squids accumulates throughout their life and is thought
y = 75x0.33, r = 0.92
to fuel an extended egg-brooding period (Seibel et al., 2000).