Edited by I. Berczi and R.M. Gorczynski 2001 Elsevier Science B.V. All rights reserved
Neuroimmune Biology — An IntroductionDepartment of Immunology, Faculty of Medicine, The University of Manitoba, Bannatyne Campus, 32-795 McDermot Avenue, Winnipeg, Manitoba R3E 0W3, Canada
That a healthy mind is fundamental to general well being has been recognized since
prehistoric times and proverbs analogous to “Healthy body – healthy mind” exist in many languages. A century ago pathologists noted fi rst that the size of the thymus was profoundly infl uenced by emotional events and by neuroendocrine aberrations. Hans Selye discovered fi rst (1936) that the hypothalamus-pituitary-adrenal axis, which is activated by diverse ‘nocuous’ stimuli, leads to the rapid involution of the thymus. He coined this phenomenon as the ‘stress’ response. Selye established that stress results in the development of the general adaptation syndrome which is characterized by elevated resistance to diverse insults. Andor Szentivanyi and colleagues discovered (1949) that hypothalamic lesions prevent anaphylactic death in guinea pigs. This is the fi rst experimental evidence for the sweeping regulatory power of the nervous system over violent, life threatening immune reactions. That the nervous system also controls the infl ammatory response was fi rst demonstrated by Milos Jancso and co-workers (1964). These fundamental discoveries were not followed by a burst of research activity. Progress has been slow because of the lack of basic knowledge and because of the immense technical diffi culties encountered.
In the seventies a handful of laboratories started to re-examine various aspects of
neuroimmune-interaction. It was established that pituitary hormones have the capacity to stimulate, inhibit and modulate immune responses. Placental and pituitary hormones are also involved in the development of the immune system and maintenance of immunocompetence. It was also described that lyphoid organs are innervated and that neurotransmitters and neuro-peptides are important immunomodulators. It became gradually apparent that immune derived cytokines and nerve impulses serve as feedback signals towards the neuroendocrine system. Compelling evidence was produced, indicating that immune reactions may be conditioned in the classical pavlovian sense and that emotions affect immune function of various organs and tissues, and in reproduction. It is also becoming obvious that Selye’s general adaptation syndrome really corresponds to the acute phase response. This is a multi-faceted and highly co-ordinated systemic defence reaction, which involves the conversion of the immune system from a specifi c, adaptive mode of reactivity to a rapidly amplifi able polyspecifi c reaction mediated by natural immune mechanisms. Immunological (poly)specifi city is assured by profoundly elevated levels of natural antibodies and liver derived proteins.
Much has been learned about the regulation of cell activation, growth and function from
immunological studies. Burnet’s clonal selectional theory designates antigen as the sole activator. Bretcher and Cohn recognised fi rst that at least 2 signals are required. This was followed by numerous studies on cell-to-cell interaction within the immune system and led to our current understanding of the importance of cell adhesion molecules and cytokines in cell activation and proliferation. This, couples with the available information about the mechanisms of action of hormones and neurotransmitters, of signal transduction and nuclear regulatory pathways paves the way to understanding how higher organisms function in their entire complexity. It is now apparent that the Nervous- Endocrine- and Immune-systems form a systemic regulatory network, which is capable of regulating all aspects of bodily functiuons in health and disease. Thus, Neuroimmune Biology provides new foundations to Biology.
Observations indicating that the central nervous system has a fundamental role in the maintenance of health has been made since prehistoric times and is referred to in proverbs of many languages. The healing power of mind and faith provides one of the important foundations of religion and is described in many religious texts. These phenomena are also observed in modern medicine and is known as the placebo effect. It has been demonstrated repeatedly by exact scientifi c methodology that patients treated with placebo in controlled medical trials do in fact show signifi cant improvement clinically in the absence of effective treatment. In ancient Persia, Egypt and in the Roman Empire fever has been regarded as a reaction with healing power. This view was maintained until modern times and during the early nineteen hundreds pyrogenic substances have been developed for the purposes of fever therapy [1-3].
About a century ago pathologists observed that acromegaly was frequently associated with
thymic hyperplasia. Hammar [4] described that the thymus frequently showed involution under the infl uence of environmental or emotional factors. In contrast, thymic hyperplasia was associated with castration, Graves’ Disease, Addison’s Disease and acromegaly. Smith described in 1930 that in hyposectomyzed (Hypox) rats the thymus regressed in weight to less than half of that of controls. In partially Hypox rats there was no involution [5].
In 1936 Hans Selye documented that the pituitary-adrenal-thymus axis was activated by
various nocuous stimuli, which led to the involution of the thymus and of the lymphoid organs [6, 7]. Moreover, Selye has established that the bursa of Fabricius in chickens was extremely sensitive to steroid hormones [8]. Within ten years Selye has proposed the theory of general adaptation syndrome (GAS) [9] on the basis of his experiments. He pointed out that this is a general reaction that leads to resistance of the organism to various insults. Selye’s scheme of GAS is shown in Figure 1, updated with current information. In 1949 Selye discovered that the infl ammatory response is regulated by corticosteroids [10]. In his article entitled “Stress and Disease” he proposed that defi cient host defense due to abnormalities of neuroendocrine factors may lead to disease [11]. Selye recognized the importance of mast cells in pathology and performed numerous studies in this respect. He summarized the knowledge about mast cell in a book [12], which is a lasting contribution on the subject.
Figure 1. Functional interrelations during the general adaptation syndrome. This fi gure is modifi ed from Selye [8]
by updating it with recent information. Solid arrows and the two broken arrows on the top with bold text is Selye’s
original fi gure. Recently identifi ed pathways are indicated with dotted arrows and with text in italics. The text below
is also from Selye. “Schematized drawing indicating that non-specifi c damage causes clinical shock, loss of body
weight and nitrogen, gastro-intestinal ulcers, temporary rise in plasma potassium with fall in plasma Cl, through
unknown pathways (nervous stimulus?, defi ciency?, toxic metabolites?) but manifestly not through the stimulation of
the hypophyseoadrenal mechanism. This is proven by the fact that the above manifestations are not prevented either
by hypophysectomy or by adrenalectomy; they even tend to be more severe in the absence of either or both of these
glands. Non-specifi c damage, again through unknown pathways, also acts upon the hypophysis and causes it to increase
corticotropic hormone production at the expense of a decreased gonadotropic, lactogenic and growth hormones. The
resulting corticotropic hormone excess causes enlargement of the adrenal cortex with signs of increased corticoid
hormone production. These corticoids in turn cause changes in the carbohydrate (sugar active corticoids) and electrolyte
metabolism (salt-active corticoids) as well as atrophy of the thymus and the other lymphatic organs. It is probable
that the cardiovascular, renal, blood pressure and arthritic changes are secondary to the disturbances in electrolyte
metabolism since their production and prevention are largely dependent upon the salt intake. The changes in γ-globulin,
on the other hand, appear to be secondary to the effect of corticoids upon the thymicrolymphatic apparatus. We do
not know as yet, whether the hypertension is secondary to the nephrosclerosis or whether it is a direct result of the
disturbance in electrolyte metabolism caused by the corticoids. Similarly, it is not quite clear, as yet, whether corticoids
destroy the circulating lymphocytes directly, or whether they infl uence the lymphocyte count merely by diminishing
lymphocyte formation in the lymphatic organs. Probably both these mechanisms are operative”.
Selye made all his contributions without knowing the function of the thymus, lymph nodes
or the bursa of Fabricius. The function of these organs was understood in the sixties and early seventies, decades after he published his seminal papers on stress. With the advent of the science of Immunology it became clear that stress has a profound immunosuppressive effect and increases the susceptibility to infectious disease. These fi ndings seemed to contradict Selye’s conclusion that the response to stress was an adaptive defense reaction which increased the resistance of the body to various noxious agents.
Andor Szentivanyi and his colleagues were the fi rst to document that the nervous system has
a dominant regulatory power over immune reactions. As a medical student Szentivanyi observed that adrenaline treatment was ineffective to alleviate an asthmatic attack in a patient. This clinical observation inspired him to do experiments in guinea pigs using anaphylactic shock as a model system. Hypothalamic lesions inhibited the development of anaphylactic shock in immunized animals [13]. Tuberal lesions (TBL) of the hypothalamus were effective in preimmunized guinea pigs and in later experiments also in rabbits to inhibit anaphylactic reactions elicited by the intravenous application of the antigen. Antibody production was also inhibited if the lesions were induced prior to immunization. The reaction of antibodies with the specifi c antigen was not affected by such lesions, nor was the release of tissue material mediating anaphylaxis. TBL temporarily increased the resistance of the animals to histamine and inhibited the anaphylactic reaction even when the animals were provided with passively transferred antibody, which elicited lethal shock in normal animals. The Schultz-Dale test, which was performed with small pieces of intestine in vitro, was also inhibited when the animals were subjected to TBL. The Arthus reaction, turpentine induced infl ammation and the Sanarelli-Schwartzmann phenomenon were unaffected by hypothalamic lesions. Lesions infl icted in other areas of the hypothalamus or the central nervous system were ineffective in modulating immune phenomena. Electrical stimulation of the mamillary region of the hypothalamus had an inhibitory effect on the anaphylactic response and increased the resistance of animals to histamine [14-16].
Szentivanyi devoted his entire career to the study of allergy and asthma. Animal experiments
pointed to the importance of the beta-adrenergic receptor in these reactions [17]. In 1968 Szentivanyi had synthesized the knowledge and all his fi ndings in a review article, entitled, “The beta-adrenergic theory of the atopic abnormality in bronchial asthma” [18]. He concluded that bronchial asthma, whether it is due to “extrinsic” or “intrinsic” causes, is ultimately elicited by the same mediators, such as histamine, serotonin, catecholamines, slow reactive substances plus cytokines. These are released during asthmatic reactions and should be considered as additional group of mediators in many tissues and in most species. Glucocorticoids are natural inhibitors of infl ammation. He proposed that the atopic abnormality in asthma is due to the abnormal function of the β-adrenergic system, irrespective of what triggered the reaction:
“The beta adrenergic theory regards asthma not as an ‘immunological disease’ but as a unique
pattern of bronchial hypersensitivity to a broad spectrum of immunological, psychic, infectious, chemical and physical stimuli. This gives to the antigen-antibody interaction the same role as that of a broad category of non-specifi c stimuli which function only to trigger the same defective homeostatic mechanism in the various specialized cells of bronchial tissue”.
Szentivanyi remained faithful to the idea of beta-adrenergic malfunction in atopy and asthma.
This is the common thread that connects the numerous papers reviews, book chapters and books he published. He studied α- and β-adrenergic receptors; adenylcyclase, cyclic-AMP and signal transduction; isolated, characterized and pharmacologically modulated phosphodiesterase; observed the systemic effect of immunization and of endotoxin on the adrenergic and cholinergic systems, on metabolism and on immune infl ammatory mediators; performed clinical studies on asthma and related conditions. His major observations were:
1. Beta-adrenergic sub-sensitivity did exist in patients with atopic dermatitis who never received
adrenergic medication. This indicates that therapeutic desensitization cannot account for the dysfunction of the beta-adrenergic system [19].
2. The beta- adrenergic reactivity of lung tissue of lymphocytes and bronchocytes from patients
with atopic asthma was found to be abnormal and various patterns of drug vs. disease-induced sub-sensitivity could be recognized [20-25].
3. Bronchial hyper-reactivity to cholinergic agents in asthma was not mediated through
cholinergic mechanisms but it was caused by the adrenergic abnormality, which was due to the so called “denervation super-sensitivity” [26-29].
4. Lymphocytes of asthmatic patients showed a signifi cant decrease in adrenaline binding to
beta-adrenergic receptors, which was independent of therapy [21, 22, 25].
Szentivanyi also studied the effects of infl ammation on β−adrenergic receptors [30-32].
In 1964 Korneva and Khai [33] described that hypothalamic lesions in commonly used
laboratory rodents (e.g. rabbits, guienea pigs, rats) inhibited the production of complement fi xing antibodies.
In 1949 Miklos Jancso and co-workers reported that capsaicin is a sensory irritant and that
repeated local or systemic administration to rats, mice and guinea pigs causes desensitization, which involves interference with pain receptors. Systemic pretreatment of animals with capsaicin or repeated local applications prevented the infl ammatory response, indicating the involvement of the nervous system. This was later confi rmed by experiments performed on denervated tissues. These observations indicated the existence of a distinct form of infl ammation, which depends on sensory nerve innervation. The stimulation of C-fi bers was necessary to induce this infl ammatory response. The neurogenic infl ammatory response was also demonstrated in man [34, 35].
It was known for some time that hormones, including those secreted by the pituitary gland,
affect immune reactions [36]. However, only after the publication of systematic studies performed on hypophysectomized rats and in animals treated with bromocriptine [37-41], was the role of pituitary hormones seriously considered in immunoregulation by the scientifi c community.
In 1975 Wannemacker and co-workers isolated the leukocyte endogenous mediator (LEM)
of fever [42], which was the fi rst immune-derived molecule identifi ed, that mediated feedback signals towards the central nervous system. Later LEM was found to be identical with interleukin-1. That IL-1 also serves as a signal for pituitary hormone release was shown by a number of investigators in the early 1980’s [43-48]. Subsequently other cytokines, especially IL2, IL6, TNF-α and interferon gamma were shown to regulate the secretion of pituitary hormones during systemic immune/infl ammatory reactions [49]. It is also clear by now that the nerves have immunoregulatory function and provide feedback signals from lymphoid organs and from sites of immune/infl ammatory reactions towards the central nervous system (CNS) [50-53].
In 1926 Metalnikov and Chorine proposed fi rst the behavioral modifi cation of the immune
response [54]. In 1933 Smith and Salinger [55] observed that asthmatic attacks were provoked in some patients with visual stimuli in the absence of the allergen. That immune reactions can be conditioned in the Pavlovian sense was demonstrated by Ader, MacQueen et al and by Gorczynski et al [56-58]. It was also observed that various cells in the immune system produce classical hormones and neurotransmitters. Smith and Blalock, Montgomery et al and DiMathia et al. [59-61] pioneered these observations.
Traditionally the cells of all tissues and organs have been divided into stromal cells, which were thought to provide for the structure of organs and the frame for the functioning cells, which were called parenchymal cells. It is now evident that stromal cells interact actively with parenchymal elements and this interaction leads to functional regulation of the tissue/organ. Moreover, invariably the stroma of all tissues and organs contain immune derived elements such as lymphocytes, macrophages or more specialized cells that include the glia cells in the nervous system, Kupffer cells in the liver, the Langerhans cells in the skin, etc. These cells contribute to function both in health and disease. Blood vessels and endothelial cells lining the blood vessels are also active participants in lymphocyte recirculation and in local immune/infl ammatory reactions. These cells interact both with the circulatory elements of the immune system and locally with elements of the tissue/organ. Cell-to-cell regulation in tissues is mediated by adhesion molecules that have complementary binding sites. These molecules are capable of delivering activation or inhibitory signals in a tissue and cell-specifi c manner [62-75].
Adhesion molecules and other cell membrane receptors have the capacity to co-aggregate
within the semi-fl uid cell membrane (capping) and allow the interaction of immunoreceptor thyrosin based activation motifs (ITAM) and -inhibitory motifs (ITIM). These motifs promote phosphorylation and dephosphoylation of signal transuding molecules, respectively. The cell may be activated or inhibited depending on the outcome of receptor interactions after capping. The relevance of these regulatory motifs to cell function is especially well established for the antigen receptors of NK cells and of T lymphocytes and for the function of Fc receptors. However, the phenomena of “receptor crosstalk” has been observed in many other systems [76-84]. These developments indicate that numerous receptors are involved in cell signaling, and that these receptors interact by multiple mechanisms that may lead to activation, inhibition or even inactivation (apoptosis) [130].
Numerous receptors in immunology and several hormone receptors need to be cross-linked
by the ligand in order to deliver an activation signal to a cell. This mechanism provides an important regulatory function in that cross linking may take place only at an optimal concentration of the ligand, whereas low or high concentrations would not be able to signal the cells. When more than one receptor isotype is available, the homo- and hetero-diamers formed by the specifi c ligand may have different regulatory functions. In addition, cross-linking may be one of the important mechanisms that promotes capping of the receptors prior to activation [76, 85, 86].
The immune system consists of mobile cells that are able to home readily to specifi c
target tissues and also to sites of infection, injury, regeneration and healing. Stromal lymphoid cells play physiological roles and are very important for host defense, regeneration and repair. Adhesion molecules mediate immunocyte homing and lymphocyte recirculation. Blood vessels also provide important barrier function in some tissues and organs that are known as immunologically privileged sites. The blood-brain barrier is very important from the point of view of neuroimmune interaction and is being extensively studied at the present time [70, 85, 87-92].
The central nervous system has the capacity to deliver neurotransmitters and neuropeptides to all tissues and cells in the body. For a long time the immune system was considered as an exception to this rule. However, it is now clear that the thymus and the spleen and other lymphoid organs are innervated. Interestingly, the spleen contains only sympathetic efferent nerve fi bers [93, 94]. Tissue mast cells are also innervated and the formation of synapses with nerve fi bers and lymphocytes can be readily demonstrated in tissue culture. Neurogenic infl ammation is the direct result of the discharge of infl ammatory mediators from mast cells after stimulation by mediators (primarily substance P) released from sensory nerve terminals. Neural mediators, such as growth factors, neurotransmitters, and neuropeptides, (e.g. substance-P, somatostatin) play major roles in the regulation of immune/infl ammatory responses. Nerve fi bers are capable of rapid and specifi c local delivery of mediators that are suitable of mounting an instantaneous reaction by initiating infl ammation. In other situations nerves may exert an anti-infl ammatory effect. The local modulation of immune reactions is equally possible by neurotransmitters and neuropeptides [93-95].
During the acute phase response there is a massive release of catecholamines into the
circulation, which is known as “sympathetic outfl ow”. Catecholamines are important regulators in the acute phase response, which is an emergency defense reaction. Sensory nerves provide feedback signals towards the CNS from sites of injury, infl ammation, and infection. The vagus nerve carries feedback signals to the CNS from visceral organs [93-96].
Historically the humoral mediators of cell-communication have been classifi ed as hormones that act at distant targets, neurotransmitters and neuropeptides, and locally produced hormone-like mediators, now called cytokines. One may also include here immunoglobulins, which originate from B-lymphocytes within the immune system. Immunoglobulins have evolved from adhesion molecules. In addition, virtually every cell membrane bound molecule is present in the serum, which includes MHC molecules and receptor-like-binding proteins. By now it is clear that “classical” hormones, neurotransmitters and neuropeptides are widely synthesized at various ectopic sites, including the immune system. Moreover, cytokines, which have been originally discovered within the immune system are now known to be synthesized in other tissues and organs, including the neuroendocrine system. Therefore, the historical defi nition of hormones, neurotransmitters and neuropeptides no longer applies. Rather, systemic and locally produced mediators complement each other, so that optimal function is assured both under physiological and pathophysiological conditions. In addition to the blood stream, lymphatic drainage of tissues, including the CNS, is important for humoral communication. The immune system receives signals from all tissues via the lymphatic system [85, 92, 97, 98].
3.1. The TRH-PRL, GH, IGF-I, TSH-thyroid axis
Thyrotropin releasing hormone (TRH) stimulates prolactin (PRL), thyroid stimulating hormone (TSH) and under some pathophysiological conditions, growth hormone (GH) release [99, 100]. Moreover, GH, PRL and TSH producing pituitary cells share the nuclear regulatory factor,
Pit-1 [101]. This suggests that these hormones represent an interdependent regulatory unit. Indeed in rats immunized with sheep red blood cells the increase of TRHmRNA was found in the hypothalamus at 4-24 hours after immunization. Pituitary TRH receptor mRNA and plasma PRL levels were also increased at the same time, while TSH and GH did not change. The hypothalamus-pituitary-adrenal (HPA) axis was activated 5-7 days after immunization. Antisense oligonucleotides complementary to TRHmRNA, given i.c.v. inhibited PRL secretion and decresed the titer of antibodies produced [102].
3.1.1. Thyrotropin releasing hormone (TRH)
TRH affects directly lymphocyte proliferation and the development of T lymphocytes in the gastrointestinal tract [103, 104]. In man, serum interleukin-2 (IL-2) levels rose signifi cantly during the standard TRH test [105]. The treatment of patients in critical illness repeatedly with TRH increased serum TSH, PRL, GH, T4 and T3 levels, and may correct the euthyroid sick syndrome [100].
3.1.2. Growth and Lactogenic Hormones (GLH)
Growth hormone, PRL and placental lactogen (PL) are referred to collectively as GLH. All three hormones show molecular heterogeneity and the variant forms of GH and PRL differ in their biological activity. GLH hormones are produced by a variety of cells in the body, including lymphocytes [106-116]. Our recent observations indicate that PRL production in lymphoid tissues is pituitary dependent (Figure 2).
GLH and cytokines (e.g. G-CSF, GM-CSF, EPO, IL-2, -3, -4, -5, -6, -7, -9, -11, -13)
share receptor structure [117-120]. Receptors for PRL and GH show heterogeneity and require cross-linking for signal delivery. At high hormone concentrations, cross-linking will not take place, but rather each receptor molecule will be bound to a separate hormone molecule, which leads to the self-inhibition of signal delivery. Homo- and heterodiamerization may take place after receptor-ligand interaction and some of the heterodiamers lead to inhibition, rather than stimulation. More than one signaling pathways play a role in GH and PRL action [86, 117, 119-123, 156]. Both GH and PRL induce the production of insulin-like growth factor-I (IGF-I) in cells of the immune system. IGF-I receptors belong to the transmembrane thyrosine kinase receptor family and are ubiquitously displayed on immunocytes [124].
The fetal pituitary gland does not play a role in the development of the immune system.
There is evidence to suggest that maternal and placental lactogenic hormones fulfi l this role [125-128]. After parturition, the function of the bone marrow, the thymus and the maintenance of immunocompetence all become pituitary dependent. The bone marrow defi ciency of hypophysectomized rats can be normalized by treatment with purifi ed PRL, GH or PL [129-131]. IGF-I plays a role in the mediation of GH action on bone marrow [132, 33]. Colony stimulating factor-1 (GM-CSF) and interleukin-3 are capable of stimulating IGF-I production in bone marrow cells and thus might function similarly to GLH in this organ [134].
GH, PRL, PL and IGF-I all stimulate thymus growth [115, 126, 135-139]. This stimulatory
effect is directly related to the maintenance of immunocompetence [136].
GH, PRL and PL all promote the antibody response [128, 140]. Human pituitary dwarf
individuals have normal immune function, which can be explained by the presence of normal serum PRL levels [141]. The dopaminergic drug, bromocriptine, suppressed humoral immunity which could be reversed by treatment with either GH or PRL. ACTH induced immunosuppression was also reversed by these hormones [142]. PRL enhanced the antibody response in mice in a biphasic manner as did syngeneic pituitary grafts [143, 144]. Cell mediated immune reactions, including contact sensitivity reactions, graft rejection, graft versus host
Figure 2. Pituitary dependence of prolactin production in lymphoid tissue. Female or male Fischer rats (150-170 g)
were used as normal controls, or were hypohysectomized (HYPOX). Some rats were treated with a rabbit antiserum
against rat prolactin (αPRL, 50 µl/day s.c.), which was initiated on day 14 after hypophysectomy, and maintained
until day 21, when extracts of organs were prepared. For the release of tissue PRL 30 mg/ml of wet tissue was
placed in serum-free RPMI 1640 culture medium and was frozen (-20oC) and thawed (37oC waterbath) three times.
The tubes were then centrifuged, and the supernatants were tested for PRL immuno- and bioactivity. Radioimmunoassy
(RIA) [359] and the Nb-2 lymphoma bioassay (BIO) [360] were used. This fi gure indicates that signifi cant quantities
of immuno- and bio-active PRL was present in the thymus and spleen of normal rats, which far exceeded serum levels.
The thymus and spleen of HYPOX rats contained only trace amounts of PRL.
reaction, and killer cell activity were stimulated by GLH [112, 128]. The tumoricidal activity of macrophages was also increased by PRL as was the cytotoxic activity of natural killer (NK) cells. High concentrations of PRL inhibited NK and lymphokine activated killer (LAK) cell activation [145]. Recombinant GH corrected the decreased NK activity in GH defi cient children [146]. GH and PRL stimulated the activities of monocyte/macropahges, and polymor-phonuclear leukocytes [147-150].
GH enhanced the production of IL-2 and IL-6 and had variable effects on IL-1 and tumor
necrosis factor-α (TNFα) production. PRL promoted IFN-γ and inhibited IL-1 production [112, 151, 152]. The age-related decline of immunocompetence may be due, at least in part, to the decline of GH/IGF-I production [134, 139, 154].
It is clear from this brief overview that GLH show redundancy as immunostimulatory
hormones. Current evidence suggests that GLH will support any function performed by the
immune system, including suppressor and killer cell activities, which is compatible with the notion of competence hormones [97].
It has been suggested on the basis of experiments performed in knockout mice, that PRL,
GH and IGF-I are not obligate immunoregulators, but rather, affect immune reactions as anabolic and stress modulating hormones [155, 156]. In actual fact the data obtained in knockout mice confi rm our original observations that GLH show overlap in the maintenance of the immune system. By no means do these knockout experiments indicate the irrelevance of GH and/or PRL to immune function. In order to prove or disprove the relevance of GLH to immunity, the entire system should be disabled. However, we predict on the basis of our observations that such mutations would have lethal consequences [157].
Much remains to be clarifi ed with regards to the role of the various isoforms of PRL and
GH, and of their receptors, in immune function. Because the receptor structure and the Jak-Stat transcription pathway of PRL and GH are shared with interleukins and hemopoietic growth factors [86, 156], some regard PRL and GH as members of the hemopoietic cytokine family. However, functional overlap with cytokines could simply indicate the capacity of systemic GLH to maintain the hemopoietic and immune systems at times when cytokines are in short supply.
TSH modulates immune function by the stimulation of thyroid hormones and also by acting on lymphoid cells. TSH receptors are expressed on dendritic cells and on CD45Rbhigh lymph node T cells. Recombinant TSH signifi cantly enhanced the phagocytic activity of dendritic cells from adult mice and selectively augmented the IL-1β and IL-12 cytokine responses following phagocytic activation. TSH also stimulated immunoglobulin secretion and IL-2 production. Human lymphocytes treated with TRH released TSH [97, 158, 159].
Thyroid hormone receptors (TR) are nuclear transcription factors and belong to the steroid-
thyroid hormone receptor family. TR is encoded by two genes, TRα and TRβ. Multiple isoforms of TR proteins are generated by alternative splicing [160]. Lymphocytes convert thyroxin (T4) to bioactive triiodothironine (T3). The effect of thyroid hormones on immune responses is variable. Enhancement, suppression, or no effect was reported repeatedly. While hypothyroidism is usually, but not always, associated with immunodefi ciency, treatment of normal animals with T3 yielded mostly negative results. In TR knockout mice (TRα-/-) thymopoiesis was suppressed. B cell maturation is depressed in mice that cannot respond to thyroid hormones [97, 155, 161].
3.2. The CRF-ACTH, α-MSH, β-END, -glucocorticoid axis
The hypothalamus-pituitary-adrenal axis (HPA) and the proopiomelanocortin (POMC) derived peptides (ACTH, α-MSH, β-END) act antagonistically to GLH and suppress adaptive immune/infl ammatory responses by acting on the nervous, endocrine and immune systems [97, 162].
3.2.1. Corticotropin releasing factor (CRF)
During acute phase immune responses, cytokines stimulate CRF, which in turn induces ACTH release. CRF integrates the stress response in the CNS and exerts a central immunosuppressive effect by the stimulation of sympathetic outfl ow. CRF is also produced within the immune system and has a direct regulatory effect on lymphocytes, which is mostly, but not always, immunosuppressive [49, 163, 164].
3.2.2. Adrenocorticotropic hormone (ACTH)
ACTH is immunosuppressive via the stimulation of glucocorticoid secretion by the adrenal gland. ACTH is produced by lymphocytes and it has a direct regulatory effect on lymphocyte proliferation, immunoglobulin production, and phagocytosis. ACTH exerts an anti-pyretic effect in the CNS [97, 162, 165, 166].
3.2.3. Beta-endorphin (β-END)β-END is produced and secreted by the pituitary gland and also within the brain and immune system. β-END and opioids in general are immunosuppressive when acting on the µ and κ opioid receptors. Opioids are also capable of immunoregulation by acting on the CNS [167-170].
4.4.4. Alpha-melanocyte stimulating hormone (α-MSH)α-MSH is a very effective antagonist of IL-1, -6, TNF, and IFN-γ. It inhibits fever and infl ammation by acting on the CNS and also exerts an antiinfl ammatory effect on peripheral targets. α-MSH promoted tolerance induction to contact sensitizing agents which was mediated by IL-10 [166, 171-173].
Glucocorticoid receptors are nuclear transcription factors and are present in all cells in the body. Glucocorticoids suppress the adaptive immune response, although evidence is increasing that basal physiological levels are actually required for the normal function of lymphocytes. Elevated pathophysiological levels (e.g. during systemic immune/infl ammatory reactions, trauma, or other stressful conditions) alter lymphocyte distribution in the body and suppress humoral and cell mediated immunity. Mononuclear and polymorphonuclear phagocyte function and cytokine production are suppressed by elevated glucocorticoid levels. On the other hand, glucocorticoids increased the expression of HLA antigens, and receptors for IFN-γ, IL-1, IL-6, Fcγ. Memory cells and the cells maintaining graft-versus-host reactions are resistant to glucocorticoids. The thymic epithelium is capable of synthesizing GC [97, 162, 174-179].
In 5 normoprolactinemic women intravenous bolus injectin of luteinizing hormone releasing hormone (LHRH) and TRH increased plasma IFN-γ levels, with the maximum response at 45 min after injection. Peak levels of PRL appeared at 15 min; TSH: 30 min; FSH: 30 min; LH: 30 min. Moreover, LHRH and TRH, separately and together, signifi cantly enhanced in vitro IFN−γ production by staphylococcal enterortoxin-A (SEA) and concanavalin (ConA)-activated peripheral blood mononuclear cells (PBMC) [180]. Luteinizing hormone (LH) has a direct stimulatory effect on the immune system. Follicle stimulating hormone (FSH) affected lymphocyte proliferation and IL-6 production [181-183]. Sex hormones play a major role in the regulation of mucosal immune responses [184].
E2 has a suppressive effect on bone marrow function, on the thymus, on T cell function, NK cytotoxicity, neutrophil and mast cell degranulation. Phagocytosis, antibody formation and certain forms of autoimmune disease are stimulated by E2. The cytotoxic activity of CD4+ cells is dependent on estrogen [97, 185-188].
Testosterone exerts a suppressive or moderating effect on the immune system, it antagonizes the enhancing effect of estrogens on various autoimmune diseases and stimulates bone marrow function. Aromatase inhibitors block the effect of testosterone on the thymus [97, 189, 190].
Dehydrotestosterone (DHT) has a stimulatory effect on T lymphocytes and immunoglobulin
formation. DHT is generated within the immune system from androstenediol or testosterone by macrophages [191, 192].
Dehydroepiandrosterone (DHEA) is a weak androgen produced in the adrenal glands. DHT
and its metabolites have emerged as major regulators of immune reactions capable of both immunostimulation and immunosuppression [193]. DHEA stimulates type 1 helper T cells (Th-1) for proliferation and IL-2 secretion and promotes cell mediated immunity. It antagonizes immunosuppression by glucocorticoids. Age related immunodefi ciency was reversed by DHEA in mice and host resistance was increased against viral, bacterial and parasitic infections. Vaccination and mucosal immunity was potentiated by DHEA. In mice DHEA administration restored the depressed splenocyte proliferation as well as IL-2, IL-3, and IFN-γ production following trauma and hemorrhage. In vitro the stimulatory effect of DHEA on splenocyte proliferation was unaltered by the testosterone receptor antagonist fl utamide, while the estrogen antagonist tamoxifen completely abrogated its effect [194]. Serum DHEA levels are decreased with aging, during chronic illness, suppressed by dexamethasone treatment, and restored by ACTH treatment [195-198].
Progesterone is a major immunosuppressive hormone and plays a key role in the harmonization of immune function with reproduction. During pregnancy, activated lymphocytes synthesize a progesterone induced blocking factor (PIBF), which inhibits NK activity and exerts an anti-abortive effect. Decidual CD56+ NK cells express PIBF. PS decreases host resistance to viral and fungal infections and inhibits the function of phagocytes [97, 199-205].
The liver produces 25-hydroxy vitamin D3, which is further processed in the kidney by 1-hydroxylase. This enzyme is also present in monocyte/macrophages, keratinocytes, bone marrow cells, placenta and in pneumocytes. The receptor for VD3 is of 50 kDa protein and belongs to the superfamily of steroid/thyroid hormone receptors [206, 207].
VD3 promotes the differentiation of macrophages, lymphocytes and of other cell types.
Monocyte/macrophage phagocytosis and cytotoxicity is promoted by VD3, whereas antigen presentation and cytokine production by T lymphocytes and cell mediated immune reactions are inhibited. NK cell mediated cytotoxicity is stimulated, B lymphocytes proliferation and immunoglobulin secretion are inhibited by VD3 treatment. Experimental autoimmune reactions are prevented by VD3 treatment. In man the treatment of psoriasis with VD3 analogues has a 100% success rate [206-213].
Melatonin (MEL) is secreted by the pineal gland. It regulates seasonal breading in animals and is involved in the regulation of circadian rhythms in vertebrates. Helper T cells express G-protein coupled MEL membrane receptors and, perhaps, MEL nuclear receptors as well.
MEL stimulates the release of Th-1 cytokines, such as IFN-γ, and IL-2, and of novel opioid cytokines which crossreact both with IL-4 and dynorphin B. MEL was found to enhance the production of IL-1, -6 and -12 in human monocytes. In general MEL exerts an immunostimulatory effect. Hematopoiesis is also infl uenced, possibly by MEL-induced-opioids acting on κ-opioid receptors that are present on stromal bone marrow macrophages. IFN−γ and colony stimulating factors appear to infl uence the production of MEL in the pineal gland. One intriguing feature of immunomodulation by MEL is that it is effective only if given at the right time within the circadian rhythm of the animal [214-220]. Much remains to be clarifi ed about MEL as an immunoregulatory factor.
Nerve growth factor (NGF) was fi rst detected in murine submandibular glands as a growth factor for sensory and sympathetic ganglia [222]. NGF belongs to the family of neurotrophins, that include brain derived neuroptrophin (BDNT) and neurotrophin-3 (NT-3). There are low affi nity neurotrophic receptors (P25) and high affi nity receptors, which are thyrosine kinases (e.g. gp140trkA for NGF; gp145trkB primarily for BNDF; gp145trkC for NT-3). Human macrophages express trkA and NGF is an autocine growth factor for these cells. The thymus, lymph nodes, express trkA and the spleen trkB, localized primarily to the stroma of these organs. There is some expression also in splenocytes and thymocytes. B lymphocytes and antigen presenting cells (follicular dendritic cells) also express receptors for NGF. NGF stimulates the growth and function of mast cells, B and T lymphocytes, stimulates IgM and IgG production, which is inhibited by IL-4. NGF inhibits the induction of IgE by IL-4 [223-237].
T and B lymphocytes, macrophages and mast cells synthesize biologically active NGF.
NGF promoted the development of hemopoietic colonies and stimulated the chemotactic and phagocytic activity of polymorphonuclear leukocytes, which suggest a proinfl ammatory role for NGF. However, in vivo the suppression of infl ammation has also been observed by NGF in several experimental models. Recent observations indicate that immune derived NGF provides protection for the nerveous system and to other host tissues during infl ammatory reactions. This phenomenon implies the existence of benefi cial ‘autoimmune’ reactions [228-249].
Leptin (LEP) is produced primarily by fat cells (adipocytes). Structurally LEP belongs to the GLH/CTK family and signals by a class I cytokine receptor (Ob-R). Two receptor isoforms are known: Ob-Ra and Ob-Rb. Leptin regulates energy metabolism, reproductive function, lymphoid development and function. Under normal physiological conditions the secretion of LEP is regulated by insulin, cortisol and sex steroids, mainly testosterone. In rats centrally administered LEP suppressed the mitogenic response of splenic lymphocytes. This was mediated through CRF-sympathetic activation. Leptin plays an important role in linking nutritional state and T cell function. In starving mice, which show immunosuppression, treatment with LEP enhanced TH1-mediated immune responses, in spite of the catabolic state of the animals. Starving animals have reduced LEP levels and show an increased sensitivity to endotoxin shock. Fasting mice respond to LPS with a blunted corticosterone and exaggerated TNF production. This could be corrected by LEP treatment [249-256].
During acute phase responses (e.g. sepsis) the serum level of LEP rises rapidly. Cytokines,
especially TNF-α, causes this elevation. LEP exerts an inhibitory effect on glucocorticoid
and IL-6 production. Blood levels of LEP correlate positively with the survival of patients with septicemia. LEP stimulates the production of IL-1 receptor antagonist (IL-1ra), which protects against LPS toxicity in mice. In murine glial cells LEP stimulated the production of IL-1β. In animal experiments exogenous LEP upregulated both phagocytosis and the production of proinfl ammatory cytokines. Leptin is also involved in wound healing and angiogenesis [252, 257-263].
3.8.1. Catecholamines and acetylcholine
Various cells in the immune system express β-type adrenergic receptors. Beta-adrenergic agents inhibit allergic and asthmatic reactions and in general inhibit various immune phenomena that include lymphoid responses to mitogens and to antigen, histamine release from leukocyctes and mast cells and skin reactions to antigen and histamine. The effect on antibody formation is variable. In vivo adrenalin elicits leukocytosis and eosinophilia which is followed by eosinopenia. At least some of these effects on leukocyte distribution are due to glucocorticoid release. Noradrenaline inhibits the histamine release from leukocytes and the degranulation of mast cells and it has a variable effect on antibody formation. In mice treated with LPS the reduction of sympathetic outfl ow by reserpine dramatically increased TNF production. Neuronal α2- and macrophage β- and α2-receptors were involved. In healthy volunteers catecholamines down regulate LPS-induced TNF-α, IL-6 and IL-1β, and increased IL 10. In patients with prolonged sepsis TNF-α and IL-6 were reduced and IL-1β and IL 10 were not modulated by catecholamines [264-273]. The role of the peripheral and central catecholamine systems on immune regulation is the subject of intense investigations at the present time.
Acetylcholine affects immune phenomena by nicotinic and muscerinic receptors. Cholinergic
agents enhance immune phenomena, including lymphocyte mitogenesis, cytotoxic reactions, the release of histamine and other mediators from mast cells. These effects are meditated by muscarine receptors. Acetylcholine stimulates the synthesis of complement components by human monocytes through the nicotinic receptor. Allergic patients show an increased sensitivity to cholinergic stimulation. The involvement of cholinergic mechanisms in exercise-induced anaphylaxis has been demonstrated [264, 274-277].
Substance-P mediates pain sensation in type C sensory nerve fi bers and is a major mediator of neurogenic infl ammation. Thymocytes, B and T lymphocytes, macrophages, mast cells and astrocytes have SP receptors. SP is capable of inducing degranulation of mucosal and intestinal type of mast cells, can cause plasma extravasation and bronchoconstriction. Substance P has a direct effect on lymphocytes, macrophages, eosinophils and neutrophils. It promotes lymphocyte proliferation, lymphokine production, and it has variable infl uence on immunoglobulin secretion. On eosinophils, SP increases Fc-γ and -ε receptors and decreases C3b receptors. SP stimulates the respiratory burst, chemotactic and phagocytic responses in polymorphonuclear leukocytes. Substance P stimulates the release of PGE and collagenase
from rheumatoid synoviocytes and of PGE and thromboxane B2 from astrocytes. Platelet cytotoxicity against Schistosoma mansoni larvae is activated by SP. SP induced IL-3 and GM-CSF production by bone marrow cells. This was partially mediated by IL-1 and IL-6, which are also induced by SP in the bone marrow. SP receptor expression is up-regulated by IL-4 and IFN-γ in murine peritoneal macrophages. The SP receptor was necessary for the normal granulomatous response to Schistosoma mansoni [278-289].
3.8.3. Calcitonin gene related peptide (CGRP)
CGRP receptors are functionally coupled to adenylate cyclase and are present on mature lymphocytes, macrophages mast cells and bone marrow cells. CGRP induces mast cell discharge, produces slow onset intense erythma in the skin and vasodilation. In human mononuclear phagocytes CGRP interferes with antigen presentation and with IFN-α induced H O production. Lymphocyte proliferation ia slso inhibited by CGRP. Nerve fi bers containing
CGRP are associated with Langerhans cells in the human skin. CGRP plays an important role in the regulation of the cutaneous immune system. It inhibited antigen presentation by human Langerhans cells, and the induction of contact hypersensitivity reactions to haptens in mice. Topically applied CGRP increased the infl ammatory response in the skin to allergens and irritants and boosted the sensitization process. In murine thymocytes CGRP inhibited the expression of NFκB and promoted apoptosis. T lymphocytes from rat thymus and mesenteric lymph nodes sythesized CGRP [286-293].
Receptors are present on T and B lymphocytes and mast cells for SOM. SOM acts as an antagonist of substance P and it has benefi cial effects in models of autoimmune disease and of chronic infl ammation. SOM inhibits IgE dependent mediator release by human basophils and mast cells. It also inhibits lymphocyte proliferation, endotoxin-induced leukocytosis, IgA secretion, IFN-α production, and affects macrophages. It has a variable effect of antibody dependent cytotoxicity [274, 275, 294-298].
3.8.5. Vasoactive intestinal peptide (VIP) and pituitary adenylate cyclase activating peptide
Receptors are present in monocytes and lymphocytes and both peptides are produced within the immune system. VIP regulates T cell homing to mucosal lymphoid tissue, it inhibits lymphocyte proliferation and has a variable effect on immunoglobulin secretion and on NK cell mediated cytotoxicity. IgA secretion was induced by VIP by isotype switching [299-303]. VIP and PACAP inhibit the nuclear translocation of NFκB in stimulated macrophages by inhibiting Jak/Stat phosphorylation and thus antagonize the effect of IFN-γ and downregulate the infl ammatory response. The production of cytokines, such as TGF-β1, IL-12, IL-4, -6, TNF-α and nitric oxide (NO) are inhibited by both peptides [304-309]. IL-6 production was enhanced by VIP/PACAP in unstimulated macrophages [310]. VIP and PACAP inhibited antigen induced apoptosis in CD4+ (but not CD8+) T lymphocytes by downregulating Fas [311]. VIP and α-MSH contribute to the immunosuppressive properties of aqueous humour in the eye [324, 25]. In man VIP inhibited the development of contact dermatitis to nickel sulphate when injected intracutaneously at the site of challenege [10].
Cytokines have been originally discovered within the immune system as humoral mediators between leukocytes (interleukins). By now it is clear that cytokines are produced in all tissues and organs in the body. Under physiological circumstances cytokines are local regulators of tissue/organ function. However, during acute phase reactions cytokines such as IL-1, TNF-α and IL6 serve as systemic hormones and induce profound neuroendocrine and metabolic alterations, which serves to boost the natural resistance of the body towards diverse noxious agents. Some other cytokines with major roles in the neuroimmunoregulatory system are interleukin-2, -4, -10, & interferon-γ (IFN-γ). Redundancy is present within the cytokine system in that
these mediators have overlapping functions. This is now well substantiated with experiments preformed in various knockout mice [97, 315-318].
These are chemotactic pro-infl ammatory mediators which are produced in response to injury, irritants, polyclonal activators, antigens and cytokines. As infl ammatory mediators CEM play important roles in host defence as well as in the pathogenesis of infl ammatory diseases. Chemokines also serve as mediators of cell-to-cell communication within the immune system and promote humoral and cell-mediated immune reactions, regulate cell adhesion, angiogenesis, leukocyte traffi cking and homing and contribute to lymphopoiesis and hematopoiesis. A vast number of CEM have been identifi ed to date which may be categorized, based on their structures, into four major groups: CXC (α), CC (β), C (γ) and CX C subfamilies. Chemokine
receptors are seven-transmembrane G proteins. Chemokines show unprecedented redundancy in receptor utilization and leukocytes express multiple receptors [315, 318]. Chemokines play a fundamental role in cell-to-cell communication throughout the body and enable every cell/tissue to emit signals towards the neuroimmune regulatory network.
The mucosal immune system consists of mesenteric lymph nodes, Peyer’s patches, the tonsils, mucosa associated lymphoid cells and lymphoid cells associated with various glands (e.g. salivary and lacrimal). It is now apparent that the mucosal immune system does not only defend the body against invading pathogens but also exerts major regulatory effects on systemic immune reactions. The problem faced by the mucosal immune system is that mucous membranes are bombarded by large amounts of antigens continuously, most of which are irrelevant to host defense. It would be counter-productive to spend a lot of immunological energy to respond to harmless antigens. On the other hand pathogenic agents and potentially harmful toxic substances must be dealt with. Immune defense is already mounted on the surface of mucous membranes, which are outside of the body. Therefore, self-non-self discrimination, which is being utilized so effi ciently by the immune system inside the body, does not apply to this situation [110, 319-323].
The initial response to antigens falling on mucosal surfaces is frequently the induction of
immunological tolerance. This may take place by clonal elimination at high antigen dosage or by active suppression if the dosage is low. Although the mechanisms that regulate tolerance induction has not been fully elucidated, it appears that antigen presentation by specialized cells that induce a distinct class of T cells capable of suppressing immune responses locally and systemically takes place. These T cells have been named as a type 3 T (TH3) cells and produce large amounts of transforming growth factor-β (TGF-β). TH3 cells exert a powerful systemic immunosuppressive effect all over the body at sites of infl ammation and on cells of lymphoid tissue [324, 325].
Mucosal mast cells are distinct from those situated in other tissues and play important roles
in the physiology and pathophysiology of mucous membranes. The submandibular gland in laboratory rodents has been identifi ed as a neuroendocrine and neuroexocrine organ secreting antimicrobial substances, immunoglobulin, hormones and enzymes that play major roles in mucosal immune reactions as well in the regulation of infl ammation, regeneration and repair within mucosal tissues and elsewhere in the body. The sympathetic superior cervical ganglion-submandibular gland axis has been suggested as one of the major immunoregulatory pathways.
The submandibular gland produces, secrets and excretes signifi cant amounts of nerve growth factor, epidermal growth factor and TGF-β, all of which are powerful immunoregulators. Glandular kallikrein, an enzyme with a potent immunosuppressive effect, is also produced. This enzyme was shown to suppress immune reactions and infl ammation when applied parenterally to animals and to play a role in the induction of oral immunological tolerance when given by mouth [322, 323, 326].
It has been demonstrated in the gut that must cells are innervated and that these cells play an
important role in intestinal absorption as well as in pathological responses, such as the initiation of infl ammation and so on. Lymphocytes exposed to antigen/infectious agents at a particular mucosal site will multiply, differentiate and redistribute to other sites of mucosal membranes through re-circulation, which provides generalized protection. This is known as the “common mucosal immune system” [285, 327].
4. BASIC CONCEPTS AND PRINCIPLES IN NEUROIMMUNOREGULATION
The rapid accumulation of experimental data in diverse systems that has relevance to neuroim-munoregulation has led to contradictions, misconceptions and confusion. These problems need to be addressed and clarifi ed. Some of the key issues are addressed below both from the theoretical and practical points of view.
It has been known for over a century that GH is capable of promoting the proportional growth of all tissues and organs in the body. Naturally, this includes the immune organs, such as the spleen, thymus and lymph nodes. Receptors for GLH are expressed on every tissue and organ in the body. Current indications are that all cells require some members of this hormone family for normal growth and function. GLH in conjunction with IGF-I exert a growth stimulatory and an anti-apoptotic effect on various cells and thus assure maintenance in a functional state, i.e. the cells are capable of responding to additional stimuli. On this basis GLH may be defi ned as competence hormones. There is much evidence in the literature indicating that the immune system is dependent on GLH for the maintenance of immunocompetence. This is related to the general function of these hormones to maintain growth and development, and lymphocyte growth is a prerequisite for adaptive immune reactions [97, 129, 135].
On the other hand, a vast amount of experimental evidence has accumulated, indicating
the role of tissue specifi c growth factors, cytokines and adhesion molecules in cell-to-cell signaling and the regulation of growth, differentiation and function of various cells in the body. It is apparent that proportional growth, although ultimately controlled by the systemic level of GH, is achieved through the coordinated interaction of systemic and local growth regulatory signals, many of which are tissue/organ specifi c and are also function-related. Current evidence indicates that GH in conjunction with IGF-I maintain all cells in a state of competence to respond to additional, function-related stimuli and produce additional regulatory mediators. Prolactin, which may be regarded as a modifi ed growth hormone, is capable of providing competence in most tissues and organs in the body with the exception of the skeleton, where its growth promoting effect is very limited. Like GH, PRL also induces IGF-I in its target cells. The IGF-I signal may be regarded as the ubiquitous cytokine signal that is needed for the survival of competent cells. On this basis one may hypothesise, that competence hormones maintain their target cells in aviable and responsive state by a direct stimulatory effect on the genome and by the induction of IGF-I secretion. Ultimately, the proliferation and functional activation of cells is determined by adhesion molecules, which are capable of delivering non-diffusible cell-to cell or cell-to-matrix signals. Adherence signals are capable of regulating cell function specifi cally on an individual basis. For example, an antigen-presenting cell delivers regulatory signals to an antigen specifi c T lymphocyte [129].
Prolactin synthesis is detectable in numerous tissues, including lymphoid tissue [97, 153].
There is compelling evidence to suggest that tissue-derived PRL fulfi ls autocrine/paracrine regulatory functions. Within the immune system, small lymphocytes are in a quiescent state and do not synthesize signifi cant amounts of mediators. They need to be activated in order to do so. Based on current experimental evidence, one may propose that inactive small lymphocytes are dependent for survival on pituitary GH/PRL and IGF-I. The dependence of cell survival in the thymus, spleen and bone marrow on GH/PRL supports this hypothesis. Moreover, pituitary PRL and GH maintain vital bodily functions and thus must act as survival hormones for the entire organism [128, 35, 328]. Animals with joint and total defi ciency of GH/PRL do not exist, and such defi ciency has not been convincingly demonstrated in man to date [154, 329]. The proposed interaction of neuroendocrine factors with adhesion signals and paracrine circuits in the regulation of bodily functions is summarized in Figure 3.
Failure of the neuroimmune regulatory system invariably leads to the death of the organism. In order to avoid frequent failures, the system must have multiple and overlapping regulatory pathways with a high degree of fl exibility and plasticity. This is achieved through isologous forms of regulatory molecules, multiple forms of receptors, and by the existence of functionally overlapping or totally interchangeable regulatory pathways. The CNS shows a high degree of plasticity. Moreover, redundancy is present in the function of growth and lactogenic hormones, of the IGF/insulin system, of steroid hormones, neuropeptides, cytokines, chemokines and of the various immunoglobin classes. It is quite common in immunology that unsuspected redundancy is revealed in the system by knocking out a particular gene. Similarly, the disabling of prolactin or growth hormone, or even IGF-I, would not paralyze immune function [85, 152]. These facts and clinical observations indicate clearly that the functional integrity of the neuroimmune regulatory network is maintained, even after very severe insults/defi ciencies due to the existence of redundant physiological and pathophysiological mechanisms.
Healthy individuals and animals maintain their body temperature, blood pressure heart rate metabolism and the concentration of various ingredients in the serum and in tissue fl uids within standard physiological ranges, which is characteristic of the species. This was fi rst recognized by Claude Bernard over a century ago, who coined the term “milieu interieur” [330], now designated as homeostasis. Under homeostatic conditions two basic forms of immune reactivity can be observed, e.g., innate or natural immunity and adaptive immunity.
The natural immune system consists of some highly specialized cells such as natural killer cells, γδ T cells and CD5+ B lymphocytes, that produce natural antibodies. The antigen
Figure 3. The interaction of neuroendocrine and autocrine/paracrine regulatory pathways. This fi gure depicts some basic
neuroendocrine and paracrine interactions in immunoregulation with reference to other cells (Please see the text for
details). It is proposed that the maintenance of all cells in the body in a functional state is dependent on competence
hormones. Additional signals are required for tissue and cell-specifi c regulation that include adhesion molecules and
cytokines. 1. Competence signal: This signal is delivered to lymphocytes and to many other cells in the body by GH
and/or PRL, produced in the pituitary gland. GH and PRL are also produced in many tissues ectopically, including
the immune system. Ectopic PRL/GH fulfi l a local regulatory function during immune reactions. It is suggested that
this autocrine/paracrine circuit makes rapid lymphocyte proliferation possible, which is a prerequisite of immune
reactions. 2.Stromal/adherence signals: Antigen presentation is best known as an activation signal for lymphocytes.
It is an adherence signal delivered by MHC molecules on antigen presenting cells. This is accompanied by additional
co-stimulatory adherence signals, which may eventually lead to lymphocyte activation. Adherence signals also play
a role in the induction of immunological tolerance, in lymphocyte survival, and in the induction of programmed cell
death (PCD). It is proposed that adhesion signals fulfi l tissue-, site-and cell-specifi c regulation in the body, i.e. the
function/fate of individual cells are determined at this level. 3.Cytokine signals: Lymphocyte activation, and cell
activation in general is completed by cytokine signals, which lead to cell proliferation, differentiation, and functional
activation. Cytokines may also perform inhibitory function (e.g. TGF-β, interferon-γ) or cause PCD (e.g. tumor necrosis
factors). a. Signal modulation: Some steroid hormones, catecholamines and endorphins/ enkephalins are capable of
modulating the process of signal delivery from the cell membrane to the nucleus by regulating Ca2+ infl ux, cAMP
and cGMP. b. Signal regulation: Thyroxin (T4) and steroid hormones (SH) control lymphocyte signaling by the
regulation of nuclear transcription factors. Thyroxin, steroid hormones and vitamin D3 play a regulatory role also in
cell differentation and in the elimination of unwanted cells via the induction of PCD. c. Local hormone activation:
Bioactive thyroid and steroid hormones are locally generated from inactive precursors by immunocytes (e.g. T3,
E2, androstenediol, androstenetriol, and vitamin D3) while the primary function of others (corticosteroids, estradiol,
progesterone, aldosterone) is systemic immunoregulation. Quiescent lymphocytes do not synthesize DNA and exert
minimum metabolic activity. Pituitary GH/PRL, some adhesion signals and serum IGF-I play a key role in the
maintenance of these lymphocytes until functional activation occurs. Neurotransmitters and neuropeptides are locally
acting functional regulators, basically acting as signal modulators and cytokines.
receptors of these cells are germ line coded, which are not subject to somatic mutation. These receptors have evolved to recognize highly conserved homologous epitopes(homotopes) on microorganisms and in self components and react instantaneously to their respective homotopes without the need of previous immunization. Natural antibodies are germ line coded and are polyspecifi c. Some liver derived proteins, namely C-reactive protein, endotoxin-binding protein, mannose binding protein, are also capable of recognizing homotopes, and to activate immune reactions [320, 331, 332].
Non-immune factors contribute to innate resistance. In this context behavioral factors,
physico-chemical factors, barriers, mucus, enzymes, anti-microbial substances, (HCL, bile acids, nitric oxide, oxygen radicals) heat shock proteins, non-immune interferons, enzymes, properdin, prostaglandins, leukotrienes, chemokines, blood clotting, species related resistance due to cell surface receptors and other factors may be mentioned. Typically it is not the antigen but cytokines and hormones that are fundamental to the regulation of natural immunity. For instance, in the regulation of NK cell mediated cytotoxicity interleukin-2, interferons, prolactin and growth hormone play important roles [332].
The adaptive immune response is initiated by antigen presenting cells that activate antigen specifi c thymus derived (T) lymphocytes bearing αβ-type antigens receptors. Bone marrow derived (B) lymphocytes recognize antigen by surface immunoglobulin molecules and produce antibodies. The antigen receptors of B and T lymphocytes undergo somatic mutation, followed by selection of those cells that do not possess self-reactivity but will react to altered self MHC antigens. Because of the elaborate selection process adaptive immune responses show exquisite specifi city. Processed antigenic fragments (epitopes) are presented to T-lymphocytes by antigen presenting cells in association with surface MHC molecules. Antigens may come from the external environment or from within the body (e.g. virus infected cells, cancer cells, autoantigens etc). Externally-derived antigens are presented by MHC class II antigens to CD4+ T cells, whereas endogenous antigens produced by virus infected and cancer cells are presented by MHC class I antigens to CD8+ T lymphocytes [85].
Although the adaptive immune response is initiated by specifi c epitopes of the antigen,
it is very well substantiated that additional “costimulatory signals” are also required for full activation of lymphocytes, for the initiation of cell proliferation, differentiation and for functional performance. In some situations the antigen signal is followed by inhibitory rather than stimulatory adherence signals, which lead to the induction of unresponsiveness, known as immunological tolerance, or anergy [85, 129].
Lymphocyte activation follows the rules of mitosis in general and involves cascades
of enzymatic reactions, which is accompanied by Ca2+ infl ux and ultimately leads to the phosphorylation of nuclear regulatory proteins. Hormones are capable of modulating the process of signal delivery from the membrane receptor to the nucleus by regulating Ca2+ infl ux, or by the modulation of cyclic nucleotide levels, or some enzymes, etc. (e.g. catecholamines, some steroid hormones, β-END and other opioid peptides) [97, 129]. This is designated as signal modulation (Figure 3). The origin of signal modulatory hormones may be exogenous or endogenous to the immune system.
Thyroid and steroid hormones, and vitamins A and D, control nuclear transcription factors
as their receptors, and are capable of regulating lymphocyte signals at the nuclear level. Because of the ability of these hormones to cross the cell membrane and home to their cytoplasmic/nuclear receptor molecules, they are capable of bypassing the cytoplasmic signal transduction pathway and amplify, suppress or even cancel completely, certain lymphocyte
signals in the nucleus. For instance, glucocorticoids are very effi cient in the inhibition of ongoing lymphocyte reactions. Hormones belonging to this category are designated as signal regulators (Figure 3). Some of these nuclear regulatory hormones (e.g. T3, E2, androstenediol and androstenetriol, vitamin D3) are synthesized within the immune system and function in an autocrine/paracrine fashion. These locally produced hormones are required for normal immune function (e.g. T3, GC, DHEA and its metabolites). Others act as powerful immunoregulators (e.g. E2, elevated GC levels, PS) and have the capacity to amplify, suppress/terminate ongoing immune reactions. Moreover, glucocorticoids are able to kill thymocytes and lymphocytes by inducing PCD, whereas other steroid hormones sensitize target cells for killer cell induced PCD [183].
The fi nal category of signals that complete the mitogenic stimulus to lymphocytes are
delivered by cytokines. If the antigenic signal is not complemented with the proper cytokine response, it is followed by activation-induced programmed cell death. The major cytokines involved in the induction of T cell mediated immunity (delayed hypersensitivity reactions, cytotoxic T lymphocytes) are IL-2 and IFN-γ. These are produced in large quantities by type 1-helper T lymphocytes (TH1). In contrast, humoral immunity is stimulated by IL-4, -5, -6 and -10, which are secreted by TH2 cells in large quantities. Although this classifi cation of T cells is very convenient, it is recognized that intermediate cells are not uncommon, which provides plasticity and redundancy in the system. The primary antibody response always starts with IgM, which is followed by switching to other immunoglobin classes (e.g. IgG, IgA and IgE) while maintaining epitope specifi city. This way a whole range of antibodies may be produced against the same epitope that have the capacity of activating diverse immune effector reactions, such as phagocytosis, complement fi xation, cytotoxicity etc., against the same target. The primary immune response needs 5-10 days to provide effi cient protection for the host, whereas secondary responses are much faster and can protect the host within less than 5 days. During the immunization period the organism must rely on natural immune mechanisms for protection [85, 332].
In health the immune system provides protection against infectious disease and diverse
insults, while homeostasis is maintained. Typically subclinical infections and insults are contained, the pathogenic agents are eliminated and the injury repaired locally. This, however, does not mean that the neuroendocrine and immune systems do not interact under these conditions. There is ample evidence to indicate that the “homeostatic milieu” with well defi ned levels of hormones, cytokines, neurotransmitters and neuropeptides is fundamental to this immune homeostasis.
Many physiological reactions can be considered as adaptive responses, which are required
for coping with altered functional demands. Thus for instance, exercise commands higher blood pressure, heart rate, altered endocrine function, metabolism, and leads to immune alterations. The hypertrophy and atrophy of organs according to functional demands is also commonly observed. Adaptive enzyme synthesis is another example of adaptive responses other than the immune response. Clearly, responses analogous to immunization, tolerance induction and apoptosis are all observable in various tissues and organs of the organism.
The highly coordinated and multi-faceted defense reaction described by Hans Selye as the general adaptation syndrome [8], is now known as the APR [334]. Fever is the unmistakable hallmark of APR, which is capable of increasing host resistant to diverse insults within hours. While liver derived proteins and natural antibodies increase rapidly during APR, the thymus
undergoes a profound involution. The adaptive immune response is suppressed. At the same time natural immune defense mechanisms are amplifi ed several hundred to a thousand times within 24–48 hours. Immune derived cytokines, primarily IL1, IL6 and TNF-α initiate the reaction by activating leukocytes and acting on the central nervous system and on numerous other organs and tissues in the body. This triggers the HPA axis for increased activity. The secretion CRF, ACTH, α-MSH, β-END and glucocorticoids is rapidly increased. Hormones of this axis suppress the adaptive immune response and regulate fever and infl ammation by acting on the nervous-, endocrine- and immune systems [324-338].
Circulating GH and PRL levels quickly rise at the beginning of febrile illness and soon
return to normal-to-subnormal levels. The IGF-I response to GH stimulation is impaired and the conversion of thyroxine (T4) to triiodothyronine (T3) in the tissues is also inhibited. Sex hormone levels are suppressed and testosterone levels may stay subnormal for lengthy periods. The levels of insulin and glucogen are consistently elevated, although insulin resistance is present [320, 331, 332, 334-338].
IL 6 levels are grossly elevated in APR. This is a pleiotropic cytokine which stimulates
the production of acute phase proteins (APP) in the liver. Glucocorticoids and catecholamines support the production of APP which rise rapidly in the serum to maximum levels (up to 1, 000 x) within 1-2 days. Natural antibody levels also show an abrupt increase. By this the serum concentration of polyspecifi c defense molecules, such as natural antibodies, LPS binding protein, C-reactive protein, mannose-binding protein is increased enormously. Complement production is also elevated, potentiating further the effi ciency of polyspecifi c defense molecules. A number of APP function as enzyme inhibitors and inhibitors of infl am-mation, which are likely to provide damage control during febrile illness [334-339]. All these changes are consistent with the rapid enhancement of polyspecifi c host resistance to infection and to various other insults as originally observed by Selye (Figure 1).
Febrile illness is an emergency defense reaction, which takes over the task of host
defense in situations when other defense mechanisms, including adaptive immunity, have failed. During APR the adaptive immune response, which is dependent on T cells, is suppressed and the immune system is placed under the command of natural antibodies and liver derived recognition molecules. These molecules are capable of recognizing homotopes on pathogens and on altered self components and activate various immune mechanisms after combining with their spcifi c target determinants. In this situation interleukin 6 is likely to function as an emergency competence hormone and insulin may be the principle growth factor fueling elevated leukocyte production and activity. Elevated serum levels of leptin ensure the energy requirements of APR. Inhibitory cytokines, such IL1 receptor antagonist, TNF synthesis inhibitor, IL10 and leukemia inhibitory factor are also elevated and participate in the regulation of infl ammatory processes. During APR INFγ excess serves as an antagonist of these cytokines [247, 255, 256, 259, 334-339].
The immunoconversion during APR from the adaptive mode of reactivity to the amplifi cation
of natural immune mechanisms provides instantaneous and rapidly increasing defense at the expense of muscles and other tissues and organs, which undergo catabolism. Therefore, the natural immune system provides the fi rst line of host defense during health and it also serves as the last resort of host defense in crisis situations. The acute phase response is a highly coordinated pathophysiological reaction where cyctokines, infl ammation and the metabolic activity of various organs and tissues are tightly regulated, all in the interest of host defense [332]. For this reason McEwen has adapted the term “allostasis” in contrast with homeostasis [340]. Indeed evidence is rapidly increasing that the “allostatic milieu” is a prerequisite for the suppression of the adaptive immune response and the amplifi cation of innate immunity.
Practical observations indicate that APR is a very effective defense reaction indeed, as in
the overwhelming majority of febrile illness recovery is the rule, which is followed by the development of specifi c immunity.
The stroma of various tissues and organsfulfi ls an important regulatory function towards the paranchymal cells that perform the specifi c tasks characteristic of the organ/tissue. Lymphoid cells (monocytes, macrophages, T and B lymphocytes, specialized antigen presenting cells, mast cells) are invariably present in the stroma and contibute to regulation. There is evidence for this in the nervous system, in the gastrointestinal tract, in the pituitary gland, in the adrenals and gonads, breast tissue and in other reproductive organs and in the skin. Cell-to-cell communication takes place within tissues and organs via adhesion molecules, which may be tissue- site- and cell-specifi c or shared with other organs and tissues. Adherence signals may promote or inhibit function, depending on the local requirements. Matrix components also deliver local regulatory signals.
PRL, GH and IGF-I maintain the cells and tissues of the body in a functional competent state.
Most tissues have the capacity to synthesize PRL, GH and IGF-I. This local production allows for tissues/organs/systems to amplify locally specifi c functions and to increase the adaptability of the organism (e.g. the adaptive immune response). Tissue specifi c growth factors may fulfi l the role of competence hormones (possible examples are: IL-2, IL-3, IL-6, GM-CSF, epidermal growth factor, fi broblast growth factor). This remains to be established. Therefore, it is suggested that tissues/organs/systems function as partially independent units, capable of generating all three categories of regulatory signals upon functional demands. This provides fl exibility and plasticity for adaptaion to the requirements that need to be fulfi lled.
Various cells of the immune system home to organs and tissues specifi cally. This is
governed by tissue specifi c adhesion molecules and by humoral signals such as chemokines, and cytokines. Monocyte/macrophage type cells and mast cells take up residence in the tissues and there is evidence for tissue specifi c differentiation. Examples of differentiation are the glia cells in the brain, mucosal versus the connective tissue type mast cells, Langerhans cells in the skin, Kupffer cells in the liver, and dendritic cells throughout the body. Endothelial cells mediate the communication between leukocytes and specifi c tissues via cytokines and adhesion molecules. The endothelium may also serve as a barrier between leukocytes and the tissue (e.g. blood brain barrier), but are also involved in the increase of vascular permeability during infl ammatory reactions.
The CNS has dominant regulatory powers in the body that includes the regulation of immune
phenomena. The regulation of the infl ammatory response by nerve impulses, the phenomenon of conditioning immune responses, the intricate and sophisticated neuroimmune mechanisms that are built into the process of reproduction in higher animals, especially mammals, the fact that emotions and stress affect immune reactivity are all examples pointing to CNS control. Immune phenomena show circadian and seasonal variation and this also indicates the existence of neuroendocrine regulatory infl uences. The sleep-wake cycle is fundamental to the maintenance of health and normal immune function [98, 285, 341-345]. Therefore the neuroimmune regulatory network is fundamental to the maintainence of health.
Our initial experiments revealed the important role of hormones secreted or regulated by the
pituitary gland in immunodefi ciency, in hematopoiesis, in the cytokine response to infectious agents, in autoimmune diseases, in host resistance to cancer and in the overall survival of
the organism [37-41, 128, 135, 154, 183, 329, 346-353]. Currently there is much evidence, indicating that abnormalities of the neuroimmunoregulatory network are associated with diseases of the nervous-, endocrine-, and immue systems and indeed, of other tissues and organs. It is beyond the scope of this overview to discuss this subject in detail. Instead, some recent publications are cited for the interested reader in addition to the ones presented in this volume [354-358].
One may conclude on the basis of available evidence that the nervous-, endocrine-, and
immune systems form a regulatory network, which is fundamental to the normal development and function of individuals from conception till death. This regulatory system also plays a role in host protection against pathological insults and in regeneration and healing. Therefore, the application of the term Neuroimmune Biology to defi ne this multi-disciplinary and integrative science is fully justifi ed.
I owe special homage to Hans Selye, who was my teacher and sparked my interest in neuroimmune interaction. Over the years many colleagues collaborated/contributed to the acquisition of the knowledge and ideas presented in this paper. I owe special thanks to Drs. Andor Szentivanyi, Eva Nagy, Henry Friesen, Kalman Kovacs, Dwight Nance, Donna Chow, Robert Shiu, Edward Baral, Richard Warrington, Lorand Bertok, John Kellen and Sylvia Asa in this respect. The experimental work discussed in this article was supported in part by MRC of Canada, The Arthritis Society of Canada, The Manitoba Health Research Council, The Manitoba Medical Services Foundation, Cancer Care Manitoba and Orion-Pharmos Corporation of Finland. I am indebted to Carol Funk and Valentina Tautkus for their devoted work on this manuscript.
Sternberg EM. The Balance Within: The Science Connecting Health and Emotions. New York: WH Freeman and Company, 2000.
Thompson WG. Placebos: A review of the placebo response. Am J Gastroenterol 2000; 95(7): 1637-43.
Nowotny A. In: Nowotny, editor. Benefi cial effects of endotoxins. New York, Plenum Press, 1993: 1-55.
Hammar JA. The new views as to the morphology of the thymus gland and their bearing on the problem of the function of the thymus. Endocrinology 1921; 5: 543-73, 731-60.
Smith PE. Effects of hypophysectomy upon the involution of the thymus in the rat. Anat Rec 1930; 47: 119-29.
Selye H. A syndrome produced by diverse nocuous agents. Nature (Lond) 1936; 138: 32.
Selye H. Thymus and adrenals in the response of the organism to injuries and intoxication. Brit J Exp Path 1936; 17: 234-248.
Selye H. Morphological changes in the fowl following chronic overdosage with various steroids. J Morph 1943; 73: 401-421.
Selye H. The general adaptation syndrome and the diseases of adaptation. J Clin Endocrinology 1946; 6: 117-230.
10. Selye H. Effect of ACTH and cortisone upon an “anaphylactoid reaction”. Can Med
11. Selye H. Stress and disease. Science 1955; 122: 625-31. 12. Selye H. The Mast Cells. Washington: Butterworth, 1965. 13. Filipp G, Szentivanyi A, Mess B. Anaphylaxis and nervous system. Acta Med Hung
14. Szentivanyi A, Filipp G. Anaphylaxis and the nervous system. Part II. Ann Allergy
15. Filipp G, Szentivanyi A. Anaphylaxis and the nervous system. Part III. Allergy 1958;
16. Szentivanyi A, Szekely J. Effect of injury to, and electrical stimulation of, hypothalamic
areas on anaphylactic and histamine shock of the guinea pig. A preliminary report. Ann Allergy 1956; 14: 259-261.
17. Townley RG, Trapani IL, Szentivanyi A. Sensitization to anaphylaxis and to some of
its pharmacological mediators by blockade of the beta adrenergic receptors. J Allergy 1967; 39(3): 177-79.
18. Szentivanyi A. The beta adrenergic theory of the atopic abnormality in bronchial asthma.
19. Szentivanyi A, Szentivanyi J, Wagner H. Measurement of numbers of adrenoceptors
in lymphocytes and lung tissue of patients with reversible obstructive airways disease. Clin Pharmacol Ther 1980; 27: 193-206.
20. Szentivanyi A. The conformational fl exibility of adrenoceptors and the constitutional basis
of atopy. Triangle 1979; 18(4): 109-15.
21. Szentivanyi A, Heim O, Schultze P. Changes in adrenoceptor densities in membranes
of lung tissue and lymphocytes from patients with atopic disease. Ann NY Acad Sci 1979; 332: 295-298.
22. Szentivanyi A. La fl exibilite de conformation des adrenocepteurs et la base constitutionelle
du terrain allergique. Rev Franc Allergol 1979; 19: 205-214.
23. Szentivanyi A, Fitzpatrick DF. The altered reactivity of the effector cells to antigenic and
pharmacological infl uences and its relation to cyclic nucleotides. II. Effector reactivities in the efferent loop of the immune response. In: Filipp G, editor. Pathomechanismmus und Pathogenese Allergischer Reaktionen. Werk-Verlag Dr. Edmund Banachewski, Grafelfi ng bei Munchen 1980: 511-580.
24. Szentivanyi A, Polson JB, Krzanowski JJ. The altered reactivity of the effector cells
to antigenic and pharmacological infl uences and its relation to cyclic nucleotides. I. Effector reactivities in the efferent loop of the immune response. In: Filipp G, editor. Pathomechanismus und Pathogenese Allergischer Reaktionen. Werk-Verlag, Dr. Edmund Banachewski, Grafelfi ng bei Munchen 1980: 460-510.
25. Szentivanyi A, Krzanowski JJ, Polson JB, Anderson WH. Evolution of research strategy
in the experimental analysis of the beta adrenergic approach to the constitutional basis of atopy. In: Oehling A, Mathov E, Glazer I, Arbesman C, editors. Advances in Allergology and Clinical Immunology. Oxford, Pergamon Press, 1980: 301-308.
26. Szentivanyi A. Effect of bacterial products and adrenergic blocking agents on allergic
reactions. In: Samter M, Talmage DW, Rose B, Sherman WB, Vaughan JH, editors. Textbook of Immunological Diseases. Boston, MA: Little, Brown and Co.1971: 356-374.
27. Szentivanyi A, Krzanowski JJ, Polson JB. The autonomic nervous system: Structure,
function, and altered effector responses. In: Middleton E, Reed CE, Ellis EF, editors. Allergy: Principles and Practice. St. Louis, MO, The CV Mosby Co, 1978: 256-300.
28. Szentivanyi A, Williams JF. The constitutional basis of atopic disease. In: Bierman
CW, Pearlman DS, editors. Allergic Diseases of Infancy, Childhood, and Adolescence. Philadelphia, PA, WB Saunders Co1980: 173-210.
29. Szentivanyi A. Adrenergic and cholinergic receptor studies in human lung and lymphocytic
membranes and their relation to bronchial hyperreactivity in asthma. In: Patient Care Publications. Darien, CT, 1982: 175-92.
30. Hackney JF, Szentivanyi A. The specifi city of glucocorticoids in the relaxation of
respiratory smooth muscle in vitro. J Allergy Clin Immunol 1975; 55: 123 (abstract).
31. Hackney JF, Szentivanyi A. The unique action of glucocorticoid succinates on respiratory
smooth muscle in vitro. The Pharmacologist 1975; 17: 271. (abstract).
32. Lowitt S, Szentivanyi A, Williams JF. Endotoxin inhibition of dexamethasone induction
of tryptophan oxygenase in suspension culture of isolated rat parenchymal cells. II. Effect of in vivo pretreatment of rats with endotoxin. Biochem Pharmacol 1982; 31: 3403-3406.
33. Szentivanyi A, Szentivanyi J. Mechanisms of action of corticosteroids. In: Proceedings of
the International Symposium on Allergy and Immunology. Lima, Peru, 1985: 531-534.
34. Korneva EA, Khai LM. Effect of destruction of hypothalamic areas on immunogenesis.
35. Jancso N. Role of the nerve terminals in the mechanism of infl ammatory reactions. Bull.
Millard Fillmore Hosp (Buffalo NY) 1960; 7-53.
36. Jancso N. Desensitization with capsaicin and related acylamides as tool for studying
the function of pain receptors. In: RKS Lim, editor. Pharmacology of Pain. Oxford, Pergamon Press, 1968.
37. Dougherty TF. Effect of hormones on lymphatic tissue. Physiol Rev 1952; 32: 379-407. 38. Nagy E, Berczi I. Immunodefi ciency in hypophysectomized rats. Acta Endocrinology
39. Berczi I, Nagy E, Kovacs K, Horvath E. Regulation of humoral immunity in rats by
pituitary hormones. Acta Endocrinology 1981; 98: 506-513.
40. Nagy E, Berczi I, Friesen HG. Regulation of immunity in rats by lactogenic and growth
hormones. Acta Endocrinology 1983; 102: 351-357.
41. Nagy E, Berczi I, Wren GE, Asa SL, Kovacs K. Immunomodulation by bromocriptine.
Immunopharmacology 1983; 6: 231-243.
42. Berczi I (author/editor). Pituitary Function and Immunity (with 12 contributing authors).
Monograph published by CRC Press, Inc., Boca Raton, FL, 1986.
43. Wannemacher RW Jr, Pekarek RS, Thompson WL, et al. A protein from polymorphonuclear
leukocytes (LEM) which affects the rate of hepatic amino acid transport and synthesis of acute-phase globulins. Endocrinology 1975; 96: 651-661.
44. Besedovsky HO, del Rey A, Sorkin E. Lymphokine-containing supernatants from Con
A-stimulated cells increase corticosterone blood levels. J Immunol 1981; 126: 385-7.
45. Berkenbosch F, van Oers J, del Rey A, Tilders F, Besedovsky H. Corticotropin-releasing
factor-producing neurons in the rat activated by interleukin-1. Science 1987; 238: 524-526.
46. Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W. Interleukin-1 stimulates the
secretion of hypothalamic corticotropin-releasing factor. Science 1987; 238: 522-524.
47. Nakamura H, Motoyoshi S, Kadokawa T. Anti-infl ammatory action of interleukin 1
through the pituitary-adrenal axis in rats. Eur J Pharmacol 1988; 141: 67-73.
48. Uehara A, Gottschall PE, Dahl RR, Arimura A. Interleukin-1 stimulates ACTH release by
an indirect action which requires endogenous corticotropin releasing factor. Endocrinology 1987; 121: 1580-1582.
49. Bernton EW, Beach JE, Holaday JW, Smallridge RC, Fein HG. Release of multiple
hormones by a direct action of interleukin-1 on pituitary cells. Science 1987; 238: 519-521.
50. McCann SM, Karanth S, Kamat A, Les Dees W, Lyson K, Gimeno M, Rettori V.
Induction by cytokines of the pattern of pituitary hormone secretion in infection. Neuroimmunomodulation 1994; 1: 2-13.
51. Pletsityi DF, Averi’anova LL. The infl uence of vagotomy on the development of
experimental endocarditis. Dokl Akad Nauk SSSR 1966; Mar 1;167(1): 238-40. Russian
52. Williams JM, Petersen RG, Shea PA, Schemdtje JF, Bauer DC, Felten DL. Sympathetic
innervation of murine thymus and spleen: evidence for a functional link between the nervous and immune systems. Brain Res Bull 1981; Jan; 6(1): 83-94.
53. Nance DM. Innervation of the spleen in the rat: evidence for absence of afferent
innervation. Burns J Brain Behav Immun 1989; Dec; 3(4): 281-90.
54. Fleshner M, Goehler LE, Hermann J, Relton JK, Maier SF, Watkins LR. Interleukin-1
beta induced corticosterone elevation and hypothalamic NE depletion is vagally mediated. Brain Res Bull 1995; 37(6): 605-10.
55. Metal’nikov S, Chorine V. Roles des refl exes conditionels dans l’immunite. Ann Inst
56. Smith GH, Salinger R. Hypersensitivity and the conditioned refl ex. Yale J Biol Med
57. Ader R. Letter: Behaviorially conditioned immunosuppression. Psychosom Med 1974,
58. MacQueen G, Marshall J, Perdue M, Siegel S, Bienenstock J. Pavlovian conditioning of
rat mucosal mast cells to secrete rat mast cell protease II. Science 1989; 243: 83-85.
59. Gorczynski RM, Macrae S, Kennedy M. Conditioned immune response associated with
allogeneic skin grafts in mice. J Immunol 1982; 129: 704-9.
60. Johnson HM, Smith EM, Torres BA, Blalock JE. Regulation of the in vitro antibody
response by neuroendocrine hormones. Proc Nat Acad Sci USA 1982; Sci 79: 4171-4.
61. Montgomery DW, Zukoski CF, Shah GN, Buckley AR, Pacholczyk T, Russell DH.
Concanavalin A-stimulated murine splenocytes produce a factor with prolactin-like bioactiv-ity and immunoreactivity. Biochem Biophys Res Commun 1987; Jun 15; 145(2): 692-8.
62. DiMattia, Gellersen B, Duckworth ML, Friesen HG. Human prolactin gene expression.
The use of an alternative noncoding exon in decidua and the IM-9-P3 lymphoblast cell line. J Biol Chem 1990; Sept 25; 265(27): 16412-21.
63. Salamonsen LA, Woolley DE. Menstruation: induction by matrix metalloproteinases and
infl ammatory cells. J Reprod Immunol 1999; Sept; 44(1-2): 1-27.
64. Kim HG, Kassis J, Souto JC, Turner T, Wells A. EGF receptor signaling in prostate
morphogenesis and tumorigenesis. Histol Histopathol 1999; Oct; 14(4): 1175-82.
65. Bodey B, Bodey B Jr, Siegel SE, Kaiser HE. Molecular biological ontogenesis of
the thymic reticulo-epithelial cell network during the organization of the cellular microenvironment. In Vivo. 1999 May-Jun; 13(3): 267-94.
66. Bianco P, Riminucci M, Kuznetsov S, Robey PG. Multipotential cells in the bone marrow
stroma: regulation in the context of organ physiology. Crit Rev Eukaryot Gene Expr1999; 9(2): 159-73.
67. Robinson GW, Karpf AB, Kratochwil K. Regulation of mammary gland development by
tissue interaction. J Mammary Gland Biol Neoplasia 1999; Jan; 4(1): 9-19.
68. Maeurer MJ, Lotze MT. Interleukin-7 (IL-7) knockout mice. Implications for lymphopoiesis
and organ-specifi c immunity. Int Rev Immunol 1998; 16(3-4): 309-22.
69. Fidler IJ. Angiogenesis and cancer metastasis. Cancer J Sci Am2000; Apr; 6 Suppl
70. King A. Uterine leukocytes and decidualization. Hum Reprod Update 2000; Jan-Feb;
71. Wagner JG, Roth RA. Neutrophil migration mechanisms, with an emphasis on the
pulmonary vasculature. Pharmacol Rev 2000; Sept; 52(3): 349-74.
72. Pirtskhalaishvili G, Nelson JB. Endothelium-derived factors as paracrine mediators of
prostate cancer progression. Prostate 2000; Jun 15; 44(1): 77-87.
73. Tanaka Y. Integrin activation by chemokines: relevance to infl ammatory adhesion cascade
during T cell migration. Histol Histopathol 2000; Oct; 15(4): 1169-76.
74. Alford D, Taylor-Papadimitriou J. Cell adhesion molecules in the normal and cancerous
mammary gland. J Mammary Gland Biol Neoplasia 1996; Apr; 1(2): 207-18.
75. Kimber SJ, Spanswick C. Blastocyst implantation: the adhesion cascade. Semin Cell Dev
76. Shiosaka S, Yoshida S. Synaptic microenvironments--structural plasticity, adhesion
molecules, proteases and their inhibitors. Neurosci Res 2000; Jun; 37(2): 85-9.
77. Renard V, Cambiaggi A, Vely F, Blery M, Olcese L, Olivero S, Bouchet M, Vivier
E. Transduction of cytotoxic signals in natural killer cells: a general model of fi ne tuning between activatory and inhibitory pathways in lymphocytes. Transplant Rev 1997; 155: 205-221.
78. Daëron M. Fc receptor biology. Annu. Rev Immunol 1997; 15: 203-34. 79. Isakov N. Immunoreceptor thyrosine-based activation motif (ITAM), a unique module
linking antigen and Fc receptors to their signaling cascades. J Leukoc Biol 1997; 61: 6-16.
80. Venters HD, Tang Q, Liu Q, VanHoy RW, Dantzer R, Kelley KW. A new mechanism of
neurodegeneration: a proinfl ammatory cytokine inhibits receptor signaling by a survival peptide. Proc Natl Acad Sci USA 1999; Aug 17; 96(17): 9879-84.
81. Yee D, Lee AV. Crosstalk between the insulin-like growth factors and estrogens in breast
cancer. J Mammary Gland Biol Neoplasia 2000; Jan; 5(1): 107-15.
82. Kawashima K, Fujii T. Extraneuronal cholinergic system in lymphocytes. Pharmacol
83. Ptasznik A, Gewirtz AM. Crosstalk between G protein-coupled receptors and tyrosine
kinase signaling: Src take centre stage. Arch Immunol Ther Exp (Warsz) 2000; 48(1): 27-30.
84. Bornfeldt KE, Krebs EG. Crosstalk between protein kinase A and growth factor receptor
signaling pathways in arterial smooth muscle. Cell Signal 1999; Jul; 11(7): 465-77.
85. Folli F, Saad MJ, Velloso L, Hansen H, Carandente O, Feener EP, Kahn CR. Crosstalk
between insulin and angiotensin II signalling systems. Exp Clin Endocrinol Diabetes 1999; 107(2): 133-9.
86. Paul WE. The immune system: An introduction. In: Paul WE, editor. Fundamental
Immunology, 4th edition. Lippincott-Raven, 1999: 1-35.
87. Clevenger CV, Freier DO, Kline JB. Prolactin receptor signal transduction in cells of the
immune system. J Endocrinol 1998; 157: 187-197.
88. Chang L, Crowston JG, Cordeiro MF, Akbar AN, Khaw PT. The role of the immune
system in conjunctival wound healing after glaucoma surgery. Surv Ophthalmol 2000; Jul-Aug; 45(1): 49-68.
89. Schwartz M, Cohen IR. Autoimmunity can benefi t self-maintenance. Immunol Today
90. Neumann H. The immunological microenvironment in the CNS: implications on neuronal
cell death and survival. J Neural Transm Suppl 2000; 59: 59-68.
91. Havran WL. A role for epithelial gammadelta T cells in tissue repair. Immunol Res
Infl ammatory mediators and modulation of blood-brain barrier permeability.
Cell Mol Neurobiol2000; Apr; 20(2): 131-47.
93. Antel J. Regulation of the Immune Response within the Central Nervous System. In:
Berczi I, Gorczynski R, editors. Neuroimmune Biology: New Foundation of Biology. Shannon, Co. Clare, Ireland, Elsevier, 2001 (in press).
94. Stevens-Felten SY, Bellinger DL. Noradrenergic and peptidergic innervation of lymphoid
organs. Chem Immunol 1997; 69: 99-131.
95. Nance D. Immunoregulation by the sympathetic nervous system. In: Berczi I, Gorczynski
R, editors. Neuroimmune Biology: New Foundation of Biology. Shannon, Co. Clare, Ireland, Elsevier, 2001 (in press).
96. McKay DM, Bienenstock J. The interaction between mast cells and nerves in the
gastrointestinal tract. Immunology Today 1994; 15: 533-538.
97. Derijk RH, Strijbos PJ, van Rooijen N, Rothwell NJ, Berkenbosch F. Fever and
thermogenesis in response to bacterial endotoxin involve macrophage-dependent mechanisms in rats. Am J Physiol 1993 Nov 265(5 Pt 2): R1179-83.
98. Berczi I, Nagy E. Immune-endocrine interactions. In: Stefaneanu L, Sasano H, Kovacs K
editors. Molecular and Cellular Endocrine Pathology. Chapman and Hall, 2000: 425-456.
99. Moldofsky H. Sleep, health and immunocompetence. In: Berczi I, Gorczynski RM,
editors. Neuroimmune Biology: New Foundation of Biology. Shannon, Co. Clare, Ireland, Elsevier, 2001 (in press).
100. Müller EE, Salerno F, Cocchi D, et al. Interaction between the thyrotropin releasing
hormone-induced growth hormone rise and dopaminergic drugs: Studies in pathological conditions in the animal and man. Clin Endocrinol 1979; 11: 645-56.
101. Van den Berghe G, de Zegher F, Vlasselaers D, Schetz M, Verwaest C, Ferdinande P,
Lauwers P. Thyrotropin-releasing hormone in critical illness: From a dopamine-dependent test to a strategy for increasing low serum triiodothyronine, prolactin, and growth hormone concentrations. Crit Care Med 1996; 24: 590-5.
102. Li S, Crenshaw EB III, Rawson EJ, Simmons DM, Swanson LW, Rosenfeld MG. Dwarf
locus mutants lacking three pituitary cell types result from mutations in the POU-domain gene pit-1. Nature 1990; 347: 528-33.
103. Castro PR, Penalva R, Pereda MP, Renner U, Reul JMHM, Stalla GK, Holsboer F, Arzt E.
Early activation of thyrotropin-releasing-hormone and prolactin plays a critical role during a T cell-dependent immune response. Endocrinology 1999;140: 690-97.
Swine Flu and the Great Flu Pandemic of 1918-19 The Similarities and What History Can Teach Us By Molly Punzo, M.D. The Great Flu Pandemic of 1918-19 killed more people than any other outbreak of disease in history. It is estimated that between 50 and 100 million died from what was then known as “Spanish Flu”. It most often killed those in the prime of life, and it killed with extra
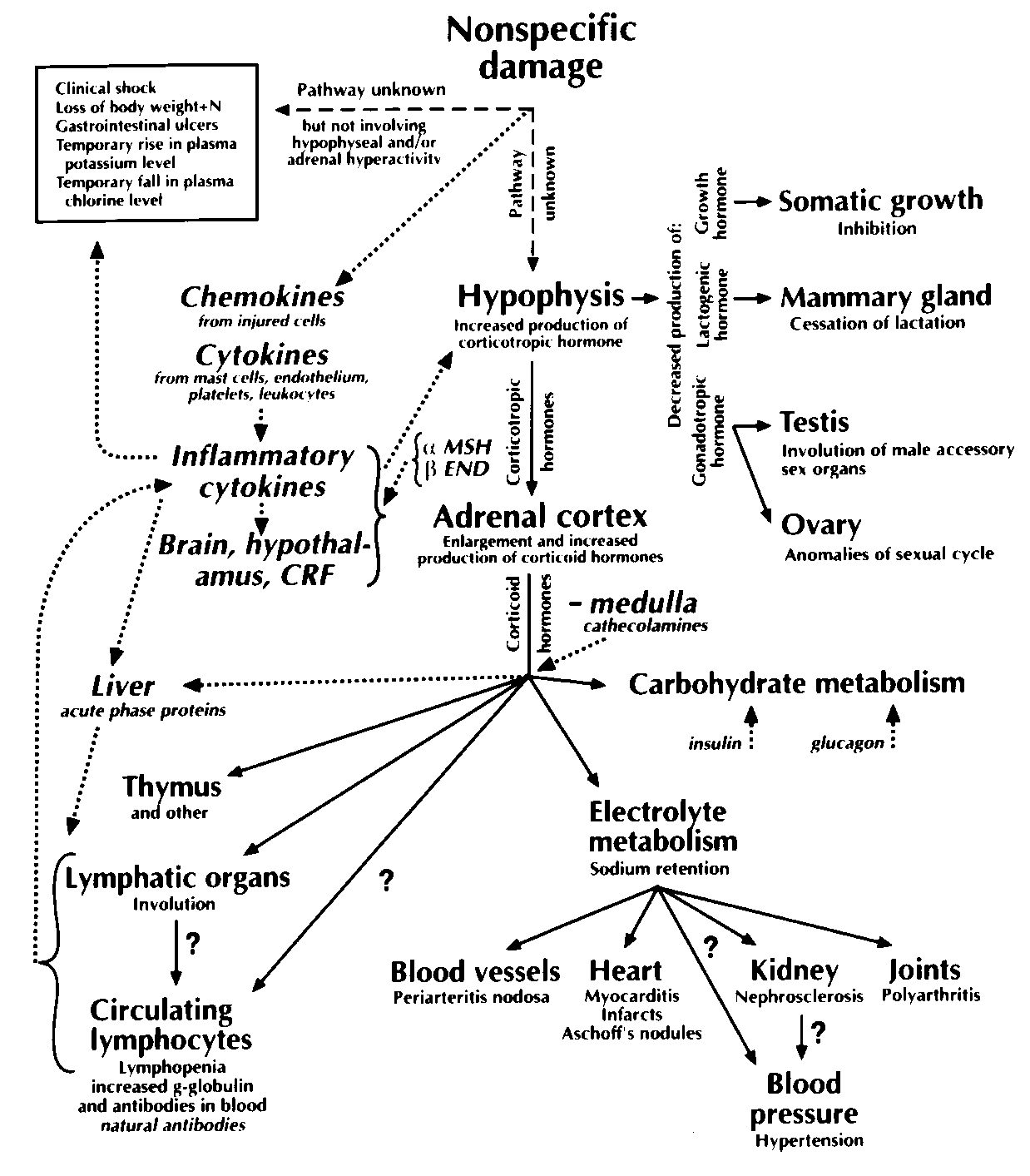 Figure 1. Functional interrelations during the general adaptation syndrome. This fi gure is modifi ed from Selye [8]
by updating it with recent information. Solid arrows and the two broken arrows on the top with bold text is Selye’s
original fi gure. Recently identifi ed pathways are indicated with dotted arrows and with text in italics. The text below
is also from Selye. “Schematized drawing indicating that non-specifi c damage causes clinical shock, loss of body
weight and nitrogen, gastro-intestinal ulcers, temporary rise in plasma potassium with fall in plasma Cl, through
unknown pathways (nervous stimulus?, defi ciency?, toxic metabolites?) but manifestly not through the stimulation of
the hypophyseoadrenal mechanism. This is proven by the fact that the above manifestations are not prevented either
by hypophysectomy or by adrenalectomy; they even tend to be more severe in the absence of either or both of these
glands. Non-specifi c damage, again through unknown pathways, also acts upon the hypophysis and causes it to increase
corticotropic hormone production at the expense of a decreased gonadotropic, lactogenic and growth hormones. The
resulting corticotropic hormone excess causes enlargement of the adrenal cortex with signs of increased corticoid
hormone production. These corticoids in turn cause changes in the carbohydrate (sugar active corticoids) and electrolyte
metabolism (salt-active corticoids) as well as atrophy of the thymus and the other lymphatic organs. It is probable
that the cardiovascular, renal, blood pressure and arthritic changes are secondary to the disturbances in electrolyte
metabolism since their production and prevention are largely dependent upon the salt intake. The changes in γ-globulin,
on the other hand, appear to be secondary to the effect of corticoids upon the thymicrolymphatic apparatus. We do
not know as yet, whether the hypertension is secondary to the nephrosclerosis or whether it is a direct result of the
disturbance in electrolyte metabolism caused by the corticoids. Similarly, it is not quite clear, as yet, whether corticoids
destroy the circulating lymphocytes directly, or whether they infl uence the lymphocyte count merely by diminishing
lymphocyte formation in the lymphatic organs. Probably both these mechanisms are operative”.
Figure 1. Functional interrelations during the general adaptation syndrome. This fi gure is modifi ed from Selye [8]
by updating it with recent information. Solid arrows and the two broken arrows on the top with bold text is Selye’s
original fi gure. Recently identifi ed pathways are indicated with dotted arrows and with text in italics. The text below
is also from Selye. “Schematized drawing indicating that non-specifi c damage causes clinical shock, loss of body
weight and nitrogen, gastro-intestinal ulcers, temporary rise in plasma potassium with fall in plasma Cl, through
unknown pathways (nervous stimulus?, defi ciency?, toxic metabolites?) but manifestly not through the stimulation of
the hypophyseoadrenal mechanism. This is proven by the fact that the above manifestations are not prevented either
by hypophysectomy or by adrenalectomy; they even tend to be more severe in the absence of either or both of these
glands. Non-specifi c damage, again through unknown pathways, also acts upon the hypophysis and causes it to increase
corticotropic hormone production at the expense of a decreased gonadotropic, lactogenic and growth hormones. The
resulting corticotropic hormone excess causes enlargement of the adrenal cortex with signs of increased corticoid
hormone production. These corticoids in turn cause changes in the carbohydrate (sugar active corticoids) and electrolyte
metabolism (salt-active corticoids) as well as atrophy of the thymus and the other lymphatic organs. It is probable
that the cardiovascular, renal, blood pressure and arthritic changes are secondary to the disturbances in electrolyte
metabolism since their production and prevention are largely dependent upon the salt intake. The changes in γ-globulin,
on the other hand, appear to be secondary to the effect of corticoids upon the thymicrolymphatic apparatus. We do
not know as yet, whether the hypertension is secondary to the nephrosclerosis or whether it is a direct result of the
disturbance in electrolyte metabolism caused by the corticoids. Similarly, it is not quite clear, as yet, whether corticoids
destroy the circulating lymphocytes directly, or whether they infl uence the lymphocyte count merely by diminishing
lymphocyte formation in the lymphatic organs. Probably both these mechanisms are operative”.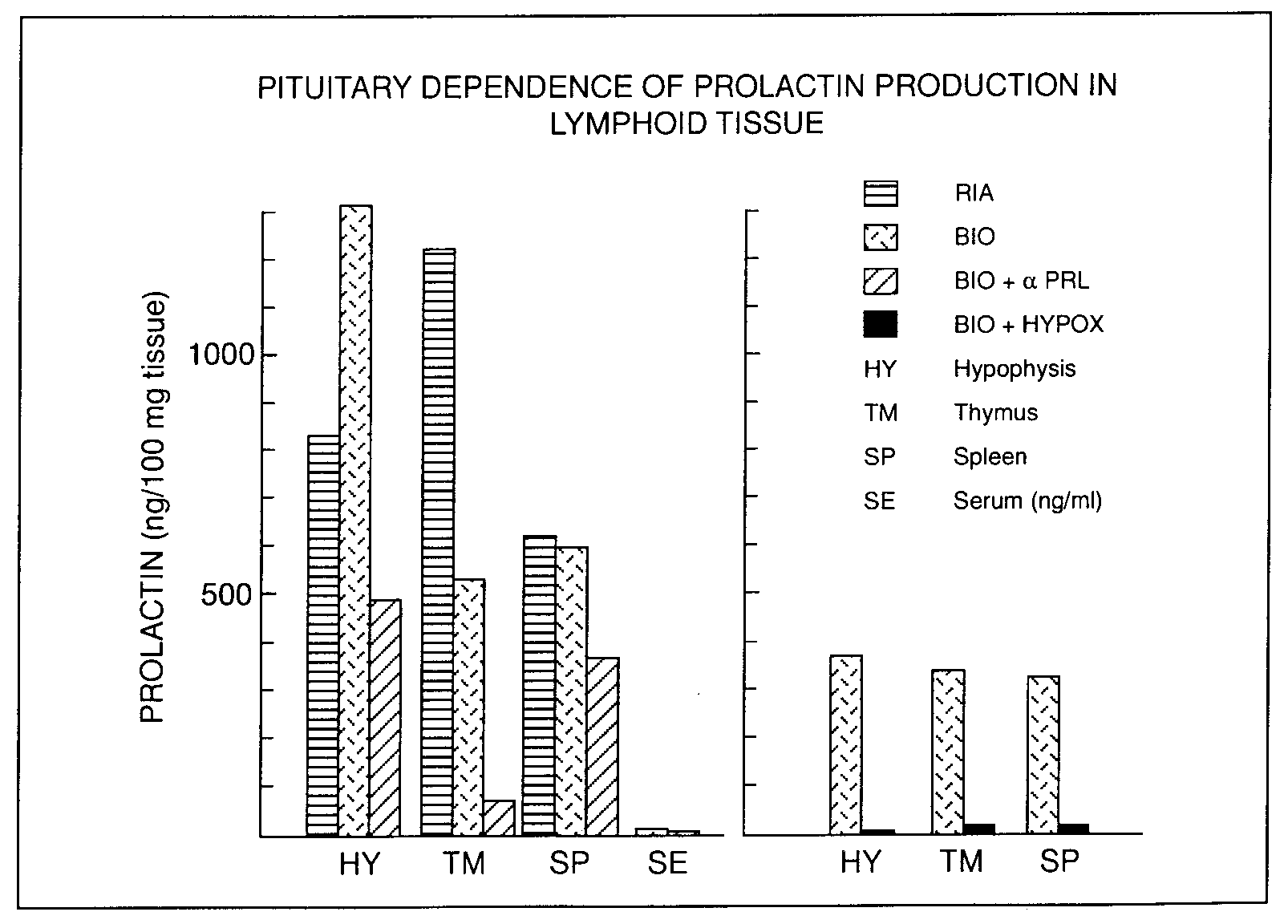 Figure 2. Pituitary dependence of prolactin production in lymphoid tissue. Female or male Fischer rats (150-170 g)
were used as normal controls, or were hypohysectomized (HYPOX). Some rats were treated with a rabbit antiserum
against rat prolactin (αPRL, 50 µl/day s.c.), which was initiated on day 14 after hypophysectomy, and maintained
until day 21, when extracts of organs were prepared. For the release of tissue PRL 30 mg/ml of wet tissue was
placed in serum-free RPMI 1640 culture medium and was frozen (-20oC) and thawed (37oC waterbath) three times.
The tubes were then centrifuged, and the supernatants were tested for PRL immuno- and bioactivity. Radioimmunoassy
(RIA) [359] and the Nb-2 lymphoma bioassay (BIO) [360] were used. This fi gure indicates that signifi cant quantities
of immuno- and bio-active PRL was present in the thymus and spleen of normal rats, which far exceeded serum levels.
The thymus and spleen of HYPOX rats contained only trace amounts of PRL.
Figure 2. Pituitary dependence of prolactin production in lymphoid tissue. Female or male Fischer rats (150-170 g)
were used as normal controls, or were hypohysectomized (HYPOX). Some rats were treated with a rabbit antiserum
against rat prolactin (αPRL, 50 µl/day s.c.), which was initiated on day 14 after hypophysectomy, and maintained
until day 21, when extracts of organs were prepared. For the release of tissue PRL 30 mg/ml of wet tissue was
placed in serum-free RPMI 1640 culture medium and was frozen (-20oC) and thawed (37oC waterbath) three times.
The tubes were then centrifuged, and the supernatants were tested for PRL immuno- and bioactivity. Radioimmunoassy
(RIA) [359] and the Nb-2 lymphoma bioassay (BIO) [360] were used. This fi gure indicates that signifi cant quantities
of immuno- and bio-active PRL was present in the thymus and spleen of normal rats, which far exceeded serum levels.
The thymus and spleen of HYPOX rats contained only trace amounts of PRL.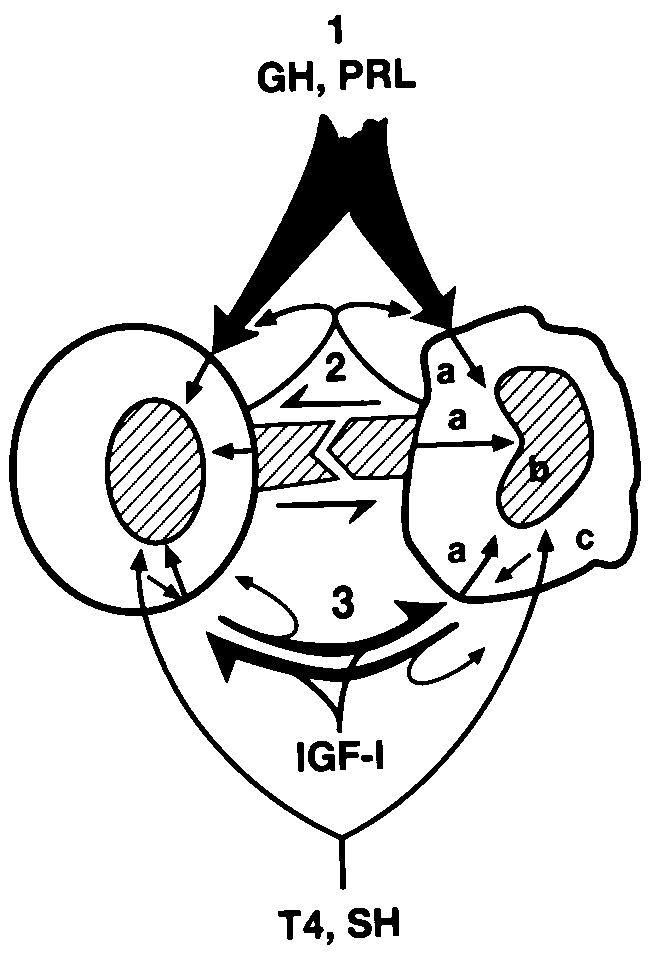 Figure 3. The interaction of neuroendocrine and autocrine/paracrine regulatory pathways. This fi gure depicts some basic
neuroendocrine and paracrine interactions in immunoregulation with reference to other cells (Please see the text for
details). It is proposed that the maintenance of all cells in the body in a functional state is dependent on competence
hormones. Additional signals are required for tissue and cell-specifi c regulation that include adhesion molecules and
cytokines. 1. Competence signal: This signal is delivered to lymphocytes and to many other cells in the body by GH
Figure 3. The interaction of neuroendocrine and autocrine/paracrine regulatory pathways. This fi gure depicts some basic
neuroendocrine and paracrine interactions in immunoregulation with reference to other cells (Please see the text for
details). It is proposed that the maintenance of all cells in the body in a functional state is dependent on competence
hormones. Additional signals are required for tissue and cell-specifi c regulation that include adhesion molecules and
cytokines. 1. Competence signal: This signal is delivered to lymphocytes and to many other cells in the body by GH