D.J. (2007a). Glutathione conjugates in the vacuole
Halkier, B.A., and Glawischnig, E. (2006).
involvement in camalexin biosynthesis through
are degraded by g-glutamyl transpeptidase GGT3
CYP71B15 (PAD3) catalyzes the final step in
conjugation of indole-3-carboxylic acid and
in Arabidopsis. Plant J. 49: 878–888.
camalexin biosynthesis. Plant Physiol. 141:
cysteine and upregulation of camalexin bio-
Parisy, V., Poinssot, B., Owsianowski, L., Buchala,
synthesis genes. J. Integr. Plant Biol. 54:
A., Glazebrook, J., and Mauch, F. (2007).
Identification of PAD2 as a g-glutamylcysteine
(2003). Functional genomics of P450s. Annu.
Weng, J.-K., Philippe, R.N., and Noel, J.P.
synthetase highlights the importance of glutathione
(2012). The rise of chemodiversity in plants.
in disease resistance of Arabidopsis. Plant J. 49:
Su, T.B., Xu, J.A., Li, Y.A., Lei, L., Zhao, L.,
Yang, H.L., Feng, J.D., Liu, G.Q., and Ren,
Winter, D., Vinegar, B., Nahal, H., Ammar,
Saga, H., Ogawa, T., Kai, K., Suzuki, H.,
D.T. (2011). Glutathione-indole-3-acetonitrile is
R., Wilson, G.V., and Provart, N.J. (2007).
required for camalexin biosynthesis in Arabi-
An “Electronic Fluorescent Pictograph”
Ohta, D. (2012). Identification and character-
dopsis thaliana. Plant Cell 23: 364–380.
browser for exploring and analyzing large-
ization of ANAC042, a transcription factor
Wada, K., Hiratake, J., Irie, M., Okada, T.,
family gene involved in the regulation of
camalexin biosynthesis in Arabidopsis. Mol.
Fukuyama, K. (2008). Crystal structures of
Zhou, N., Tootle, T.L., and Glazebrook, J.
Plant Microbe Interact. 25: 684–696. Escherichia coli g-glutamyltranspeptidase in
(1999). Arabidopsis PAD3, a gene required for
Saito, K., Hirai, M.Y., and Yonekura-Sakakibara,
complex with azaserine and acivicin: novel
camalexin biosynthesis, encodes a putative
K. (2008). Decoding genes with coexpression
mechanistic implication for inhibition by gluta-
cytochrome P450 monooxygenase. Plant Cell
networks and metabolomics - ‘Majority report by
mine antagonists. J. Mol. Biol. 380: 361–372.
precogs’. Trends Plant Sci. 13: 36–43.
Wang, M.-Y., Liu, X.-T., Chen, Y., Xu, X.-J., Yu,
Ziegler, J., and Facchini, P.J. (2008). Alkaloid
Schuhegger, R., Nafisi, M., Mansourova, M.,
B., Zhang, S.-Q., Li, Q., and He, Z.-H. (2012).
biosynthesis: Metabolism and trafficking. Annu. Arabidopsis acetyl-amido synthetase GH3.5
Reply: Complexity in Camalexin Biosynthesis
accumulation of multiple glutathione S-
camalexin pathway in Arabidopsis.
subunits of GGTs from bacteria and animal.
cides with camalexin production;GSTF6
overexpression increased and GSTF6
et al., 2011) may reflect inhibition of GGPs
only 4.8 to 9.8% sequence identities with
Arabidopsis thaliana GSTF6 expressed in
acivicin has been widely used as a specific
yeast cells catalyzed GSH(IAN) formation;
inhibitor of GGT activity for in vitro and in
conserved residues involving in the inter-
action of GGTs with acivicin are present in
Arabidopsis GGTs, but not GGPs. There-
biosynthetic pathway; and, finally, g-glu-
binding of acivicin with Arabidopsis GGTs
acid residues in substrate binding pocket.
inhibits GGT activity. However, experimen-
data and the expression patterns of these
GGTs from bacteria and animal cells (Wada
hypothesis of Møldrup et al. (2013) that
genes, we suggest that (1) GSH is the Cys
et al., 2008). For example, Escherichia coli
GGT binds acivicin through the formation of
inhibits GGP’s activities. For example, we
thesis, (2) the conjugation of GSH with IAN
a covalent bond between the Og atom of its
binding with the GGPs, the binding affinity,
dihydroisoxazole ring of acivicin and also
and/or the crystal structure of GGP-acivicin
camalexin biosynthesis. The Letter to the
alternative explanation for reduced cama-
Flores et al., 2011) argue that g-glutamyl
lexin production following acivicin treat-
2008). To show if Arabidopsis GGTs and
ment, suggesting that “termination of all
GGT activity could arrest glutathione and
www.plantcell.org/cgi/doi/10.1105/tpc.113.109975
glutathione conjugates in the vacuole and
Figure 1. Multiple Sequence Alignment of Arabidopsis GGT1, GGT2, GGP1, GGP3, and GGTs from Other Organisms.
The coding sequences of all genes were aligned by ClustalW (MegAlign program DNAStar). Sequences corresponding to the S-subunits of GGTs frombacteria and animals are presented. Identical residues in all GGTs are boxed. The residues that interact with acivicin in E. coli GGT are indicated by anasterisk above the alignment.
extracellular space” (Møldrup et al., 2013).
in the camalexin secretion. Most recently,
porters in Arabidopsis, are reported to be
Møldrup et al. (2013) also questioned the
uole and cytosol for GSTF6 (Carter et al.,
our experiments (Su et al., 2011). They note
lexin biosynthetic pathway to be cytosolic
based on the cytosolic localization of some
after inhibition of GGT activity or elimination
known enzymes in the pathway. Their point
2011; Møldrup et al., 2013) will be difficult
of GGT gene expression is possibly due to
of view is that enzymes in a biosynthetic
pathway usually are colocalized temporally
secretion. In fact, the liquid media system
and spatially (Møldrup et al., 2013). Be-
camalexin biosynthetic pathway. It is a gen-
erally accepted viewpoint that enzymes in
a biosynthetic pathway might be located in
et al., 2008; Su et al., 2011) and by other
groups (Bednarek et al., 2005; Ren et al.,
ing to this viewpoint, the different localiza-
as described by Xu et al. (2008). The camalexin inliquid media and GSH(IAN) in seedlings weremeasured as described (Su et al., 2011).
Sequence data from this article can be found in theArabidopsis Genome Initiative or GenBank/EMBLdatabases under the following accession numbers:MKK9, At1g73500; GGT1, At4g39640; GGT2,At4g39650; Escherichia coli GGT, NP_417904;Helicobacter pylori GGT, NP_207909; humanGGT, NM_005265; pig GGT, NM_214030; ratGGT,
GGP3, At4g30550. T-DNA insertion lines usedhere are as follows: ggt1-1 (SAIL_1178_C01) andggt2-1 (SAIL_6_G02).
Figure 2. Camalexin and GSH(IAN) production in MKK9DD, Columbia Wild-Type, ggt1-1, and ggt2-1Seedlings.
Twelve-day-old seedlings were either treated with dexamethasone (DEX) or inoculated with Botrytiscinerea spores. Twelve hours after dexamethasone treatment or 16 h after inoculation with B. cinerea,the seedlings were used for GSH(IAN) extraction and the media were used for camalexin measurement.
This work was supported by grants from the
Right axis, peak areas for GSH(IAN) (mass-to-charge ratio [M1H]1 ¼ 462.17); left axis, camalexin
National Natural Science Foundation of China
content. Data represent the means 6 SD of three biological replicates. Asterisks indicate statistically
(No. 31030010 and 31125006 to D.R., No.
significant differences between wild-type (WT) and GGT mutant seedlings. **P , 0.01 (paired sample t
31000127 to Y.L., and No. 30771124 to H.Y.).
likelihood that a given enzyme is involved in
a particular pathway in planta,” as noted by
T.S., Y.L., and H.Y. performed the experiments.
T.S., Y.L., H.Y., and D.R. wrote the article.
colocalization cannot be taken as evidence
that such enzymes do not work in the same
camalexin than the wild type after patho-
Another observation that led Møldrup et al.
(2013) to argue that GGPs instead of GGTs
are involving in camalexin biosynthesis is that
(IAN) in GGT mutants in our previous work
ported that GH3.5, a multifunctional acetyl-
(Su et al., 2011). However, we have since
amido synthetase, is involved in camalexin
measured the contents of GSH(IAN) in MKK9
biosynthesis through catalyzing the conju-
hyperactive mutants (as MKK9DD), Columbia
gation of indole-3-carboxylic acid with Cys.
camalexin biosynthesis. We found that GSH
biosynthetic pathway. Current data there-
(IAN) accumulated in the wild type and in
Badri, D.V., Chaparro, J.M., Manter, D.K.,
MKK9DD, ggt1, and ggt2 mutants undergoing
Martinoia, E., and Vivanco, J.M. (2012).
camalexin biosynthesis. Figure 2 shows that
GSH(IAN) accumulation is positively corre-
chosen by Arabidopsis plants likely de-
porters in root exudation of phytoalexins,
lated with camalexin production. The result
signals, and in disease resistance. Front. Plant
substrate [e.g., GSH(IAN)] in order to produce
Oldham, N.J., and Hahlbrock, K. (2005).
As shown in Figure 2 of Møldrup et al.
Structural complexity, differential response toinfection, and tissue specificity of indolic and
(2013), both the ggt1 and ggt2 mutants
Arabidopsis roots. Plant Physiol. 138: 1058–
wild type after AgNO3 treatment. However,
Arabidopsis thaliana wild type (ecotype Columbia-
Carter, C., Pan, S., Zouhar, J., Avila, E.L.,
biological repeats in their experiments, the
0), ggt1-1 and ggt2-1 mutants, and MKK9DD
authors suggest that there are no signifi-
transgenic seedlings were all grown and treated
vegetative vacuole proteome of Arabidopsisthaliana reveals predicted and unexpected
B.A. (2013). Assigning gene function in bio-
Su, T., Xu, J., Li, Y., Lei, L., Zhao, L., Yang, H.,
proteins. Plant Cell 16: 3285–3303.
synthetic pathways: Camalexin and beyond.
Feng, J., Liu, G., and Ren, D. (2011).
Geu-Flores, F., Møldrup, M.E., Bo¨ttcher, C.,
Glutathione-indole-3-acetonitrile is required
Olsen, C.E., Scheel, D., and Halkier, B.A.
Nafisi, M., Goregaoker, S., Botanga, C.J.,
for camalexin biosynthesis in Arabidopsis
(2011). Cytosolic g-glutamyl peptidases pro-
Glawischnig, E., Olsen, C.E., Halkier, B.A.,
thaliana. Plant Cell 23: 364–380.
cess glutathione conjugates in the biosynthesis
and Glazebrook, J. (2007). Arabidopsis cyto-
Wada, K., Hiratake, J., Irie, M., Okada, T.,
of glucosinolates and camalexin in Arabidopsis.
lyzes the conversion of indole-3-acetaldoxime
Fukuyama, K. (2008). Crystal structures of
Hull, A.K., Vij, R., and Celenza, J.L. (2000).
in camalexin synthesis. Plant Cell 19: 2039–
Escherichia coli g-glutamyltranspeptidase in
Arabidopsis cytochrome P450s that catalyze
complex with azaserine and acivicin: Novel
the first step of tryptophan-dependent indole-
Ohkama-Ohtsu, N., Radwan, S., Peterson, A.,
mechanistic implication for inhibition by gluta-
3-acetic acid biosynthesis. Proc. Natl. Acad.
Zhao, P., Badr, A.F., Xiang, C., and Oliver,
mine antagonists. J. Mol. Biol. 380: 361–372.
D.J. (2007). Characterization of the extracel-
Wang, M.-Y., Liu, X.-T., Chen, Y., Xu, X.-J., Yu,
Mao, G., Meng, X., Liu, Y., Zheng, Z., Chen, Z.,
lular g-glutamyl transpeptidases, GGT1 and
B., Zhang, S.-Q., Li, Q., and He, Z.-H. (2012).
and Zhang, S. (2011). Phosphorylation of a
GGT2, in Arabidopsis. Plant J. 49: 865–877. Arabidopsis acetyl-amido synthetase GH3.5
WRKY transcription factor by two pathogen-
Ren, D., Liu, Y., Yang, K.-Y., Han, L., Mao, G.,
involvement in camalexin biosynthesis through
synthesis in Arabidopsis. Plant Cell 23:
conjugation of indole-3-carboxylic acid and
phytoalexin biosynthesis in Arabidopsis. Proc.
cysteine and upregulation of camalexin bio-
Martin, M.N., Saladores, P.H., Lambert, E.,
Natl. Acad. Sci. USA 105: 5638–5643.
synthesis genes. J. Integr. Plant Biol. 54:
Hudson, A.O., and Leustek, T. (2007). Local-
Schuhegger, R., Nafisi, M., Mansourova, M.,
ization of members of the g-glutamyl trans-
Xu, J., Li, Y., Wang, Y., Liu, H., Lei, L., Yang,
peptidase family identifies sites of glutathione
Halkier, B.A., and Glawischnig, E. (2006).
H., Liu, G., and Ren, D. (2008). Activation of
and glutathione S-conjugate hydrolysis. Plant
CYP71B15 (PAD3) catalyzes the final step in
MAPK kinase 9 induces ethylene and camalexin
camalexin biosynthesis. Plant Physiol. 141:
biosynthesis and enhances sensitivity to salt stress
Møldrup, M.E., Geu-Flores, F., and Halkier,
in Arabidopsis. J. Biol. Chem. 283: 26996–27006. Reply: Complexity in Camalexin Biosynthesis
Tongbing Su, Yuan Li, Hailian Yang and Dongtao Ren
; originally published online February 28, 2013;
This information is current as of December 13, 2013
References
This article cites 16 articles, 12 of which can be accessed free at:
Permissions CiteTrack Alerts Subscription Information
e t of the Georgian National Platform of the Eastern Partnership’s conference held on 15 September 2012 “Parliamentary elections and Georgia’s EU integration prospects”: The 1 October parliamentary election is an extremely important challenge for Georgia’s statehood and democratic development. Ensuring a free and fair election environment and holding the elections to high standard
CENIFAR MENDOZA, 02 DE MAYO DEL 2013 Artículo Nombre GTIN 96288 ACLASTA 5 MG/100ML F.A. X 1 07795306045659103549 ACTEMRA 200MG/10ML VIAL X 1 07792371933867103550 ACTEMRA 400MG/20ML VIAL X 1 0779237193388188646 AGRELID 0,5MG CAP X 100 0779534916898888647 AGRELID 1MG CAP X 100 0779534916904628537 ANTIBIOPTAL COL X 5 ML 0779536800066589198 BARACLUDE 0,5MG COM X 30 0300003161122789199 BA
 Figure 1. Multiple Sequence Alignment of Arabidopsis GGT1, GGT2, GGP1, GGP3, and GGTs from Other Organisms.
Figure 1. Multiple Sequence Alignment of Arabidopsis GGT1, GGT2, GGP1, GGP3, and GGTs from Other Organisms.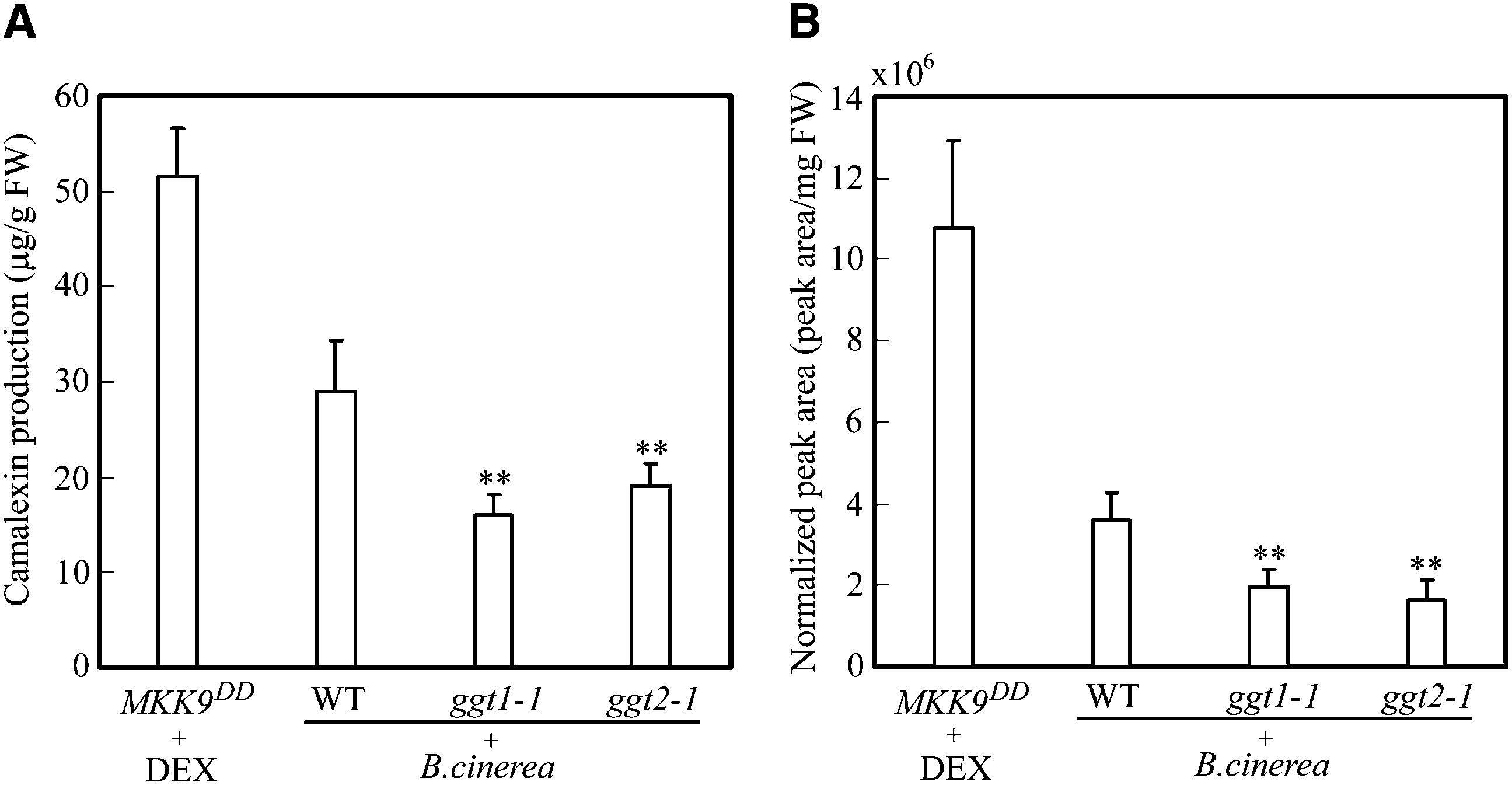 as described by Xu et al. (2008). The camalexin inliquid media and GSH(IAN) in seedlings weremeasured as described (Su et al., 2011).
as described by Xu et al. (2008). The camalexin inliquid media and GSH(IAN) in seedlings weremeasured as described (Su et al., 2011).