Blood pressure and heart rate in the ovine fetus:ontogenic changes and effects of fetal adrenalectomy
NOBUYA UNNO,1,2 CHI H. WONG,1,3 SUSAN L. JENKINS,1 RICHARD A. WENTWORTH,1XIU-YING DING,1 CUN LI,1 STEVEN S. ROBERTSON,4 WILLIAM P. SMOTHERMAN,3AND PETER W. NATHANIELSZ11Laboratory for Pregnancy and Newborn Research, Department of Physiology, College of VeterinaryMedicine, and 4Department of Human Development, Cornell University,Ithaca, New York 14853-6401; 3Laboratory of Perinatal Neuroethology, Department of Psychology,Binghamton University, Binghamton, New York 13902; and 2Department of Obstetricsand Gynecology, Faculty of Medicine, University of Tokyo, 113 Tokyo, JapanUnno, Nobuya, Chi H. Wong, Susan L. Jenkins, Rich-
weeks of gestation as a result of both an increase in
ard A. Wentworth, Xiu-Ying Ding, Cun Li, Steven S.
cardiac output and a rise in peripheral vascular resis-
Robertson, William P. Smotherman, and Peter W.
tance (18), whereas the decrease in FHR has been
Nathanielsz. Blood pressure and heart rate in the ovine
ascribed to a baroreflex response to the increased FABP
fetus: ontogenic changes and effects of fetal adrenalectomy.
(19), resulting in increased parasympathetic influence
Am. J. Physiol. 276 (Heart Circ. Physiol. 45): H248–H256,1999.—Ontogenic changes in baseline and 24-h rhythms of
via the vagus on basal FHR (48). However, the exact
fetal arterial blood pressure (FABP) and heart rate (FHR) and
mechanisms responsible for these changes are un-
their regulation by the fetal adrenal were studied in 18 fetal
known, partly because ontogenic changes in FABP and
sheep chronically instrumented at 109–114 days gestation
FHR have not been fully characterized.
(GA). In the long-term study, FABP and FHR were continu-
It has been shown that influences of -sympathetic
ously recorded from 120 days GA to spontaneous term labor
and parasympathetic activity on baseline FHR increase
(Ͼ145 days GA) in five animals. Peak times (PT) and ampli-
with gestational age in the sheep fetus (47). In addition,
tudes (Amp) of cosinor analysis were compared at 120–126,
plasma concentrations of hormones that have stimula-
127–133, and 134–140 days GA. Consistent, significantlinear increases in FABP and linear decreases in FHR were
tory effects on the fetal cardiovascular system increase
observed in all fetuses. Significant 24-h rhythms in FABP and
with gestational age (25, 34, 41). However, relative
FHR were observed during all the time windows. In the
roles of the fetal endocrine and autonomic nerve system
adrenalectomy study, to test the hypothesis that fetal cortisol
in the ontogenic changes in fetal cardiovascular system
plays a key role in cardiovascular maturation, fetal adrenals
have not been characterized. Because glucocorticoids
were removed in eight animals (ADX); sham fetal adrenalec-
have a pronounced stimulatory effect on blood pressure
tomy was performed on five animals (Con). Cortisol (4 µg/
(BP) in adult (40) and fetal (11, 12, 45, 50) sheep, it is
min) was infused intravenously in four ADX fetuses from day
possible that the ontogenic changes in the fetal cardio-
7 postsurgery for 7 days (ADXϩF). No significant changes in
vascular system are, at least in part, regulated by the
PT and Amp in FABP and FHR were observed. Plasmacortisol levels remained low in Con and ADX fetuses (Ͻ4.9
fetal hypothalamic-pituitary-adrenal axis. In one previ-
ng/ml). Cortisol infusion increased fetal plasma cortisol to
ous study, bilateral adrenalectomy (ADX) in fetal sheep
22.3 Ϯ 3.2 ng/ml (mean Ϯ SE) on day 13 in ADXϩF fetuses.
at 119 to 133 days of gestation (GA) produced no
FABP increased in control and ADXϩF but not ADX fetuses;
significant changes in FHR and FABP compared with
FHR decreased in control and ADX but rose in ADXϩF
intact fetuses (35). In a second study, short-term (5 h)
fetuses. These results suggest that, in chronically instru-
intrafetal cortisol infusion at Ͼ132 days GA to intact
mented fetal sheep at late gestation, 1) increases in FABP
fetuses produced an increase in plasma cortisol concen-
and decreases in FHR are maintained consistently from 120
tration to 6.3 Ϯ 0.7 ng/ml and caused a transient
to 140 days GA, with distinct 24-h rhythms, the PT and Ampof which remain unchanged, and 2) the physiological increase
increase in FABP and a decrease in FHR (50). Finally,
in FABP is dependent on the fetal adrenal; bilateral removal
continuous cortisol infusion at a rate of 4 µg/min to
of the fetal adrenals does not prevent the ability of cortisol to
intact fetuses induced a sustained increase in FABP for
produce a sustained increase in FABP.
up to 48 h when administered at 103–120 days GA, but
cardiovascular system; circadian rhythm; adrenal; cortisol
had no effect at 130–137 days GA (12, 45). Thesestudies demonstrated that glucocorticoids can act toincrease FABP, although they are not required for themaintenance of basal FABP. In addition they showed
IN THE SHEEP FETUS during late gestation, arterial blood
that the effect of glucocorticoids on FABP is gestational
pressure (FABP) increases steadily (6, 10, 22) and fetal
age dependent. In adult sheep, extensive investigations
heart rate (FHR) declines steadily (22). It has been
have been conducted on the BP increases produced by
postulated that the FABP increases during the last few
both ACTH and cortisol. It has been shown that ACTHadministration induces an increase in cardiac outputwith a consequent rise in BP within 24 h unaccompa-
The costs of publication of this article were defrayed in part by the
nied by changes in peripheral vascular resistance (40).
payment of page charges. The article must therefore be hereby
In adult sheep it has also been reported that the
marked ‘ advertisement’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
ACTH-induced BP increase is not abolished by treat-
0363-6135/99 $5.00 Copyright 1999 the American Physiological Society
ment with ␣- and -adrenergic blockade, angiotensin-
converting enzyme inhibitors, or ganglion blockades
Surgery was performed under halothane general anesthe-
(43), suggesting that mechanisms in addition to the
sia on five ewes between 113 and 114 days GA in the
sympathoadrenomedullary and the renin-angiotensin
long-term study and on 13 ewes between 109 and 113 days GA
system may be involved in mediating the hypertensive
in the ADX study using techniques that have been described
in detail (30, 31, 36). Briefly, polyvinyl catheters were in-
Short-term cortisol infusion in intact fetal sheep at
serted into a maternal carotid artery and jugular vein and
Ͼ132 days GA decreases plasma norepinephrine and advanced into the arch of the aorta and superior vena cava,epinephrine concentrations (50). This suppression of
respectively. The uterus was then exposed through a midline
the fetal sympathoadrenomedullary system suggests
Long-term study. Hysterotomy was performed, and fetuses
that catecholamines play a relatively unimportant role
were instrumented with polyvinyl vascular catheters in-
in the cortisol-induced BP increase. However, more
serted via the carotid artery and jugular vein. Multistranded
direct experiments to explore the roles of catechol-
stainless steel wire (Cooner Sales, Chastsworth, CA, catalog
amines in the maintenance of the elevated BP have not
no. AS 632) electrodes were sewn to the myometrium.
been conducted. Inasmuch as a previous study demon-
ADX study. Hysterotomy was performed. In eight fetal
strated that cortisol stimulated epinephrine release
sheep both fetal adrenal glands were exposed and isolated via
from cultured fetal adrenal medulla cells (17), it is
a retroperitoneal approach and removed. In five fetuses the
possible that the adrenal medulla plays a role in the
adrenals were exposed but not removed (Con; n ϭ 5) (30). Fetuses were instrumented with polyvinyl catheters inserted
maintenance of the cortisol-induced FABP increase.
via the femoral artery and tibial vein. An amniotic cavity
Furthermore, no information is available that ad-
dresses the effects of fetal ADX on the ontogenic
After surgical preparation of the ewe and fetus, all fetal
changes in FHR or the effects of prolonged elevation of
catheters and leads were grouped to exit the lateral abdomi-
plasma cortisol levels on basal FHR.
nal wall of the ewe at a single point. Surgical closure was
Several studies have demonstrated the existence of
accomplished in layers. The ewe returned to the laboratory.
24-h rhythms in FABP (7) and FHR in sheep (7, 9, 24).
During the four days after surgery, the ewe received 1 g/day iv
However, no study to date has examined whether there
ampicillin sodium. The ewes were fed daily, and water was
are ontogenic changes in amplitudes and peak times of
available ad libitum. All fetuses were allowed to recover for atleast 5 days after surgery before being studied.
the 24-h rhythms in these fetal cardiovascular vari-
Maternal and fetal arterial blood samples (0.5 ml) were
ables. It has been suggested that glucocorticoids play a
taken daily after the surgery for measurements of blood gases
significant role in the regulation of 24-h rhythms of
and pH on a blood gas analyzer (ABL500, Radiometer,
FHR in the human fetus (1); however, there is no
Copenhagen, Denmark). Measurements were corrected to
information on the changes in the 24-h rhythms of
39°C. A heparin solution (10 U/ml physiological saline) was
FABP and FHR after fetal ADX or sustained premature
continuously infused at a rate of 0.5 ml/h into each vascular
increases in plasma cortisol concentrations in the sheep
catheter to ensure that catheters remained open.
In the present study we studied the chronically
instrumented sheep fetus to test the hypothesis that
From the sixth day after the surgery, FABP, FHR, and
fetal cortisol plays a key role in cardiovascular matura-
myometrial electrical activities were recorded continuously
tion in late gestation. We characterized 1) the ontogenic
throughout the study with the use of a data acquisition
changes in FABP and FHR and 2) the ontogenic
system that collected data averaged every second (16). FABP
changes in the amplitude and the peak time of 24-h
and amniotic cavity pressure were measured continuously
rhythms in these fetal cardiovascular variables by
with the use of a calibrated pressure transducer (Cobe,
measuring hourly FABP and FHR continuously be-
Lakewood, CO) connected to the fetal artery and amnioticcavity catheters. Amniotic fluid pressure was taken as the
tween 120 and 140 days GA. In addition we also
zero pressure reference for FABP. FHR was calculated from
investigated 1) the effects of fetal ADX on the ontogenic
the BP systolic peak to peak intervals. In each fetus, averaged
changes in FABP and FHR and 2) the effects of fetal
values of FABP and FHR were calculated every 40 s and
ADX on cortisol-induced increases in FABP between
analyzed with an IBM compatible personal computer using
Microsoft Excel. Inappropriate signals due to blood sam-plings, fetal movements, and catheter malfunctioning wereexcluded. Hourly and daily averaged values for FABP and
MATERIALS AND METHODS
FHR were calculated beginning at 0000 (Eastern Standard
Time) on 120 days GA in the long-term study and at 1600 onthe sixth day after surgery in the ADX study.
Eighteen Rambouillet-Columbia crossbred ewes bred on a
single occasion only and carrying a fetus of known gestational
age were used. All procedures were approved by the CornellUniversity Animal Care and Use Committee. All facilities
Long-term study. After the onset of labor, confirmed by the
were approved by the American Association for the Accredita-
presence of irreversible contraction-type myometrial electri-
tion of Laboratory Animal Care. From 7 days before surgery,
cal activities, the ewe and the fetus were killed with an
the ewe was housed in a metabolic stall with ad libitum
overdose of pentobarbital sodium (Fatal-Plus, Vortech Phar-
alfalfa cubes and water in a room with controlled light-dark
maceuticals, Dearborn, MI) and the body weight was deter-
cycles (lights on at 0700 and off at 2100). ADX study. From six days after surgery, 5 ml of maternal
ADX study. Fetal body and organ weights were compared
and fetal blood were collected daily between 0900 and 1000,
among the three treatment groups using one-way ANOVA.
and plasma was removed, frozen in liquid N2, and stored at
Changes in daily values of fetal blood gases and pH were
Ϫ20°C until assayed for ACTH and cortisol. ADX fetuses were
analyzed among the three groups using one-way ANOVA and
divided into two groups. Four adrenalectomized fetuses
in each treatment group using one-way RM ANOVA. Data for
(ADXϩF) were continuously infused with cortisol (Solu-
fetal plasma ACTH and cortisol concentrations were com-
Cortef, Upjohn, Kalamazoo, MI) via the fetal venous catheter
pared among the three treatment groups by one-way ANOVA
at a rate of 4 µg/min starting at 1600 on the seventh day after
or by Kruskal-Wallis nonparametric ANOVA where appropri-
surgery until necropsy. The four other adrenalectomized
ate. Changes in daily FABP and FHR values were compared
fetuses (ADX) received vehicle alone. Fetuses were delivered
by one-way RM ANOVA in Con and ADX fetuses. In ADXϩF
by cesarean section and killed by exsanguination while under
fetuses, daily FABP and FHR were analyzed between the
halothane general anesthesia at 123–125 days GA. Complete-
values on the day before infusion (day Ϫ1), 1 day after the
ness of ADX was confirmed in all ADX and ADXϩF fetuses by
commencement of infusion (day 1), and the fifth day of
careful inspection of the surgical sites. Tissues were collected
infusion (day 5) with the use of one-way RM ANOVA. Post hoc
analyses for multiple comparisons were performed with theSNK test. Cosinor analysis was carried out to determine the
presence of 24-h rhythms and to evaluate any change pro-
Plasma ACTH concentrations were measured with a com-
duced by ADX and ADXϩF. Cosinor curves were fitted to the
mercial RIA kit (INCStar, Stillwater, MN) validated for
hourly FABP and FHR data averaged in each animal from
hormone measurements in sheep plasma (46). Assay sensitiv-
day 1 to day 6. Peak times and amplitudes were compared
over three treatment groups by one-way ANOVA.
coefficients of variation (CV) for quality control samples
For all statistical tests, differences were considered to be
containing 34.7 (pool of the assay kit), 10.9 (fetal pool), and
53.9 (maternal pool) pg/ml were 6.8 and 12.5%, 12.8 and
19.0%, and 6.5 and 10.7%, respectively. Plasma cortisolconcentrations were measured with a commercially available
RIA kit (Diagnostic Products, Los Angeles, CA) validated formeasurements in sheep plasma (46). Intra-assay CV was
Spontaneous labor was confirmed by the presence of
8.8% for a quality control sample containing 36.1 ng/ml (n ϭ
contraction-type myometrial electrical activity at
20). Interassay CV was 2.3% for a quality control sample
146.7 Ϯ 0.5 days GA. At necropsy, fetal body weight was
containing 29.9 ng/ml (n ϭ 20). Assay sensitivity (90% B/B0)
Arterial blood gases and pH in ewes and fetuses.
Mean values for arterial blood gases and pHa in ewesand fetuses are presented in Table 1. There were no
All data are presented as means Ϯ SE. Data were analyzed
significant changes in pHa and arterial blood gases.
first by the summary of measures method (27) to focus the
Overall changes in FABP and FHR. After we deleted
unusable, corrupt, or unavailable periods of recording
Long-term study. Daily average values for FABP and FHR
signals, hourly values were obtained in 95 Ϯ 2% of total
were determined in each fetus. Changes in FABP and FHRwith gestational age were analyzed with one-way repeated-
period in each fetus. No cardiovascular data were
measures (RM) ANOVA followed by the Student-Newman-
analyzed within 2 days of labor. Daily average values
Keuls (SNK) test for post hoc comparisons. Cosinor analysis
for FABP and FHR from 120 to 143 days GA are
was performed on hourly average values in each animal to
presented in Fig. 1. FABP increased steadily with
determine the presence of 24-h rhythms (33). Cosinor curves
gestational age from 35.2 Ϯ 1.7 mmHg on 120 days GA
were fitted to the hourly FABP and FHR data in each animal
to 47.4 Ϯ 2.4 mmHg on 143 days GA, whereas FHR
over 7-day periods (120–126, 127–133, and 134–140 days
decreased with gestational age from 178 Ϯ 3 beats/min
GA) after the subtraction of the linear component based on
on 120 days GA to 143 Ϯ 2 beats/min on 140 days GA.
the results of the linear regression analysis. Among animals
Between 140 and 143 days GA baseline FHR increased.
that revealed a significant 24-h rhythm, peak times and
Ontogenic changes in 24-h rhythms in FABP and
amplitudes were compared among three groups using one-way RM ANOVA. For the statistical analysis of arterial blood
FHR. Hourly values in FABP and FHR beginning at
gases and arterial pH (pHa), one-way RM ANOVA was
2400 on 120 days GA until 2300 on 143 days GA are
applied on weekly average values, which was followed by the
illustrated in Fig. 2. Cosinor analysis on 24-h rhythms
of cardiovascular variables revealed significant 24-h
Table 1. Arterial blood gases and pH in ewes and fetuses in the long-term study
Data are presented as means Ϯ SE of 5 animals. Blood samples were taken at 0900–1000 every day beginning at 120 days gestation (GA).
Data were averaged in each animal to obtain representative values for each period. pHa, PaCO , Pa , arterial pH, PCO
No significant differences were observed throughout the study period.
Table 2. Peak time amplitude of 24-h rhythms in FABPand FHR in the long-term study
Values are means Ϯ SE of 5 fetuses, except in fetal arterial blood
pressure (FABP) at 120–126 days GA and 127–133 days GA, wheren ϭ 4. FHR, fetal heart rate. No significant differences were observedthroughout the study period. Lights were on at 0700 and off at 2100.
Fig. 1. Daily fetal arterial blood pressure (FABP) and daily fetalheart rate (FHR) (n ϭ 5 ewes). Values were presented as means Ϯ SE.
pHa values in ADXϩF fetuses increased significantly
FABP increased from 120 to 143 days gestation (GA), whereas FHR
after cortisol infusion. Arterial partial pressure of O2
decreased steadily from 120 to 140 days GA. bpm, Beats/min.
(PaO ) in ADX fetuses was significantly lower than Con
fetuses between the sixth and ninth day after surgery.
rhythms in FABP in four of the five fetuses between
This difference in PaO was not observed during the rest
120–126 and 127–133 days GA and five fetuses during
of the experimental period. A serial analysis revealed a
134–140 days GA; FHR showed significant 24-h
significant increase of PaO in ADXϩF fetuses after
rhythms in all of the five fetuses during all three
gestational age windows. Peak times and amplitudes of
Plasma ACTH and cortisol concentrations. In Con
24-h variations in FABP and FHR remained unchanged
fetuses, fetal plasma ACTH levels remained stable
throughout the study period (Table 2).
throughout the observed period (22.2 Ϯ 3.3, 20.6 Ϯ 2.4,and 22.2 Ϯ 3.3 pg/ml on day Ϫ1, day 3, and day 6,
respectively). Plasma ACTH concentrations in ADX
Fetal body and organ weights. At necropsy ADX
fetuses were significantly greater compared with those
fetuses and ADXϩF fetuses did not show any signifi-
in Con fetuses (113 Ϯ 36, 107 Ϯ 31, and 128 Ϯ 37 pg/ml
cant difference in body weight or in the weight of the
on day Ϫ1, day 3, and day 6, respectively). In the
fetal heart, lungs, kidneys, liver, spleen, or thymus
ADXϩF group, the plasma ACTH concentration on day
compared with Con fetuses (Table 3).
Ϫ1 was 120 Ϯ 93 pg/ml. After cortisol infusion, plasma
Fetal blood gases and pH. pHa and arterial partial
ACTH concentrations decreased and reached a level of
14.6 Ϯ 2.5 pg/ml on day 3. Fetal plasma ACTH re-
out the experimental period, although pHa values in
mained low (15.1 Ϯ 1.8 pg/ml on day 6) during the rest
ADX fetuses were significantly lower than those in Con
fetuses on the eighth and tenth day after surgery and
There was no significant difference in maternal
plasma cortisol concentration among the three groups,except on day 1, when the ADXϩF mothers showedsignificantly higher cortisol concentrations comparedwith Con and ADX mothers. No consistent interrelation-ship was found between maternal and fetal plasmacortisol concentrations. In Con and ADX fetuses, plasma
Table 3. Fetal body and organ weights
Fig. 2. Hourly FABP (A) and hourly FHR (B; n ϭ 5) beginning at
Values are means Ϯ SE in grams; n ϭ 4 ewes each group. Con,
0000 on 120 days GA until 2300 on 143 days GA. Values are presented
control; ADX, adrenalectomized; ADX ϩ F, ADX and cortisol-infused
Table 4. Arterial blood gases and pH for fetuses of Con,Baselines of FABP and FHR. Daily baseline FABP
ADX, and ADX ϩ F in the ADX study
and FHR values on the sixth day after surgery were39.6 Ϯ 1.4 and 190 Ϯ 2, 39.4 Ϯ 1.6 and 185 Ϯ 1, and
44.9 Ϯ 1.9 mmHg and 187 Ϯ 3 beats/min in Con, ADX,
and ADXϩF fetuses, respectively. There were no differ-
ences among these groups. Hourly FABP and FHR
values are shown for Con, ADX, and ADXϩF fetuses in
Changes in FABP and FHR in Con and ADX. During
the study period there was a significant increase in
FABP associated with a significant decrease in FHR in
Con fetuses. In contrast there was no significant change
in FABP in ADX fetuses, whereas the decrease in FHR
was similar to those in Con fetuses.
ADXϩF fetuses increased significantly on the first dayof cortisol infusion compared with values of the preced-
ing day (Fig. 4C). This increase was sustained through-
out the study period. FHR also increased after cortisol
infusion (Fig. 4F). This increase was gradual compared
with the FABP increase and remained sustained
throughout the infusion period, which reached a signifi-
cant level on the fifth day after the commencement of
Changes in 24-h rhythms of FABP and FHR in ADXand ADXϩF. Cosinor analysis revealed a significant
24-h rhythm in FHR in three of four Con fetuses, four of
four ADX fetuses, and four of four ADXϩF fetuses. No
Values given are means Ϯ SE; numbers in parentheses are number
difference was found in the peak time or amplitude
of fetuses. Significant differences: * ADX vs. Con; † ADX or Con vs.
among the three treatment groups. The peak time and
ADX ϩ F (1-way ANOVA, P Ͻ 0.05).
amplitude of each treatment group were not signifi-cantly different from those of the fetuses in the long-
cortisol concentrations remained below the assay sensi-
term study. Cosinor analysis was also applied to FABP,
tivity level for the volume of plasma extracted (4.9
and a significant 24-h rhythm was found in two of four
ng/ml) throughout the observation period. In ADXϩF
Con fetuses, four of four ADX fetuses, and three of four
fetuses, plasma cortisol concentrations increased signifi-
cantly after the commencement of cortisol infusion andremained ϳ23 ng/ml (Fig. 3). DISCUSSION Long-term study. This study is the first to report
measurement of FABP and FHR continuously over thecritical period of development from 120 days GA todelivery in the sheep fetus. This approach enableddetailed analysis on ontogenic changes in baselineFABP and FHR and in their 24-h rhythms. Kitanaka etal. (22) reported a steady increase in FABP and asimultaneous decrease in FHR from 110 to 120 days GAover 21 days in the sheep fetus. Because they measuredFABP and FHR for only 1 h every day, it was difficult todetect small differences in the trajectory of developmen-tal changes in these parameters. Brace and Moore (7)found that both FABP and FHR have 24-h rhythms inthe late gestation sheep fetus, although they did notspecify the gestational ages at which the study was
Fig. 3. Fetal plasma cortisol concentrations in adrenalectomized,cortisol-infused (ADXϩF) fetuses. Values are means Ϯ SE (n ϭ 4).
conducted. In the present study, we used well-accli-
Dotted line indicates sensitivity of the assay. * On the sixth and
mated sheep and computer-based data acquisition us-
seventh day after surgery, all values were below the sensitivity of the
ing carefully calibrated transducers and amplifiers to
assay. Cortisol infusion to fetuses at a rate of 4 µg/min was com-
achieve longitudinal continuous recording of FABP and
menced after the blood sampling on the seventh day after surgery
FHR for 24 days. We demonstrated clear and consistent
and continued until necropsy. Values in control and ADX fetuses werebelow the sensitivity of the assay throughout.
ontogenic changes in FABP and FHR from 120 to 143
Fig. 4. Hourly FABP and hourly FHR. Values aremeans Ϯ SE beginning on the sixth day (i.e., 24-hbefore the beginning of cortisol and/or vehicleinfusion) in control (Con), ADX, and ADXϩFfetuses. Each day starts at 1600. A: hourly FABPin Con fetuses (n ϭ 5). B: hourly FABP in ADXfetuses (n ϭ 4). C: hourly FABP in ADXϩFfetuses (n ϭ 4); D: hourly FHR in Con fetuses(n ϭ 4); E: hourly FHR in ADX fetuses (n ϭ 3); F:hourly FHR in ADXϩF fetuses (n ϭ 3). Barindicates period of cortisol infusion in C and F. * First day of significant sustained increase fromday Ϫ1.
days GA with distinct 24-h rhythms. These findings
decrease in PaO is related to the FABP profile in ADX
strengthen previous studies and provide important
fetuses, because the FABP profile did not change after
information to understand the mechanisms of the
the recovery of PaO during the latter one-half of the
ontogeny of the fetal cardiovascular system.
experiment. Further studies are required to evaluate
ADX study. To examine the possible roles of the fetal
the precise causal mechanism and overall physiological
adrenals in the changes in FABP and FHR that have
relevance of the effect of ADX on the rise in FABP that
been characterized in the long-term study, we investi-
occurs at this stage of gestation. Cortisol infusion to
gated the effects of fetal ADX on the normal gestational
ADX fetuses beginning at 117 days GA resulted in a
age-related changes in FABP and FHR and on the
significant increase in FABP that was similar to previ-
previously described increase in FABP produced by
ous findings in intact fetal sheep (12, 45). This increase
infusion of glucocorticoids to the fetus (12, 45, 50). A
in FABP was sustained for Ͼ6 days throughout the
recent study reported a significant reduction in fetal
cortisol infusion period. These results clearly indicate
body weight 4 wk after fetal ADX at 111–114 days GA
that the fetal adrenal medulla does not play an indis-
(49). It is possible that long-term changes in fetal
pensable role in mediating cortisol-induced FABP in-
conditions after fetal ADX affect not only fetal growth
creases in late gestation fetal sheep. A previous study
but also ontogenic changes in the fetal cardiovascular
on adult sheep reported that total autonomic blockade
system. Therefore, in the present study we evaluated
does not attenuate ACTH-induced increases in FABP
the effect of ADX for 2 wk after fetal ADX at the criticalperiod of adrenal development. At necropsy, no differ-
(43), supporting our conclusion that the adrenal me-
ences were observed in fetal body and organ weights
dulla is not critically involved. However, these observa-
among Con, ADX, and ADXϩF fetuses, which supports
tions do not exclude the possibility of interaction of
the concept that overall fetal condition was substan-
glucocorticoids at either the receptor or postreceptor
tially unchanged in all animals during the study pe-
level with locally released catecholamines. The sus-
riod. In a previous study we demonstrated that the
tained effects of cortisol on FABP for up to 6 days in the
average fetal plasma cortisol over the 5 days before
present study support and extend the results of a
spontaneous vaginal delivery in control fetuses was
previous study of the effects of 48-h cortisol infusion to
59 Ϯ 10 ng/ml (28). Thus the levels of replacement we
intact fetuses in which the FABP increase after cortisol
achieved (Ϫ23 ng/ml) were within the physiological
infusion was evaluated for 48 h (12). Our findings also
range the fetus reaches in late gestation.
suggest that the cortisol-induced increase in FABP is
BLOOD PRESSURE. After ADX, the FABP increase that
not transient but may involve a fundamental change in
normally occurs with gestation was attenuated, suggest-
the regulation of the fetal cardiovascular system.
ing a significant contribution of the fetal adrenals to the
The chronic hypertensive effect of glucocorticoids
gestational age-related BP increase in fetal sheep.
during development we and others have demonstrated
Although PaO values in ADX fetuses were significantly
may play a role in the more long-term effects on BP that
lower than Con fetuses during the first one-half of the
follow prenatal glucocorticoid exposure demonstrated
experimental period, it is not likely that this temporary
in rats (3). Because growth retardation has been linked
with development of high BP later in life (2) and an
FHR decrease, such as baroreflexes, before circulating
increase in fetal plasma cortisol concentrations in
cortisol starts to increase exponentially at ϳ140
cordocentesis samples obtained from growth-retarded
human fetuses has been reported (13), this stimulatory
Twenty-four-hour rhythms in fetal cardiovascular
action of cortisol on the fetal cardiovascular system
system. Synchronized diurnal variations in BP and
could be involved in the mechanism of adult hyperten-
heart rate exist in the adult in many species, including
sion and/or cardiovascular diseases of fetal origin. In
rats (42), rabbits (14, 38), marmosets (39), monkeys
addition, our findings of maintained effects on BP over
(15), and humans (26), peak times of which correspond
6 days also have relevance to possible consequences of
to the active period for respective species. In the sheep
repeated antenatal glucocorticoid therapy adminis-
fetus, similar diurnal rhythms in FABP and FHR have
tered to women in threatened premature delivery over
been described (7). However, ontogenic changes in the
cardiovascular diurnal rhythms during fetal life have
FETAL HEART RATE. Because the gestational age-
not been characterized. Results of the present study
related FHR changes are unaffected by ADX, our
support previous findings and further indicate the
findings suggest an insignificant role of fetal adrenal
absence of ontogenic changes in the 24-h rhythms in
maturation in this aspect of cardiac function. Changes
FABP and FHR between 120 and 140 days GA in the
in FHR in ADXϩF fetuses indicate a stimulatory effect
sheep fetus. The mechanisms responsible for the 24-h
of sustained elevation of plasma cortisol on basal FHR
rhythms in the cardiovascular system have not been
despite the concurrent increase in FABP. Because
fully identified. It has been shown that the suprachias-
baroreflexes are present and functional in the late
matic nucleus, which is known as a ‘‘biological clock’’ in
gestation sheep fetus (5), this stimulatory effect could
mammals (23), plays a role in this phenomenon (20,
be a result of alteration of the setting of the baroreflex
37). There is substantial evidence that suggests that
responses. However, this is unlikely because changes in
diurnal rhythms in BP and heart rate are under
FHR in ADXϩF fetuses were completely opposite to
sympathetic control (4, 21). A previous study in the
FHR changes in Con and ADX fetuses. Alternatively, in
human fetus reported that the 24-h FHR rhythm
rats, it has been demonstrated that glucocorticoids
disappears after maternal and fetal adrenal gland
increase postsynaptic sensitivity of the cardiovascular
suppression with triamcinolone (1), suggesting the
system to norepinephrine (8). It has been also reported
adrenocortical regulation of fetal 24-h rhythms. The
that glucocorticoids enhance the sensitivity of the
lack of changes in amplitudes of the 24-h rhythm in
pacemaker -adrenergic receptors to catecholamines
FHR observed in the present study may suggest that
(29). In adult sheep, Spence et al. (43) reported that
the 24-h rhythm in the fetal cardiovascular system is
acute ganglion blockade increased FHR to a greater
regulated by an independent factor from fetal develop-
level in ACTH hypertensive sheep than in normoten-
ment, such as maternal endocrine environment, and/or
sive controls and suggested that ACTH treatment may
that the fetal mechanisms for this phenomenon are
have a direct chronotropic action on the heart. Glucocor-
already established at 120 days GA. Furthermore, the
ticoids have also been suggested to play a key role in
lack of an effect of fetal ADX with or without subse-
the developmental changes in the function of cardiac
quent continuous cortisol supplementation on the 24-h
-adrenergic receptors in the rat (32). In the sheep rhythm of FHR suggests the lack of involvement of bothfetus, a recent study (44) demonstrated that intrafetal
fetal adrenal cortical and medullary effects on the fetal
cortisol infusion at a rate of 0.5 mg · kgϪ1 · hϪ1 for 60 h to
24-h rhythm. Both the human and sheep data would be
fetal sheep at 128 days GA produced no changes in
compatible with a role for the maternal but not the fetal
myocardial -adrenergic receptor density and affinity;
adrenal in regulating these rhythms. Further studies
however a significant increase in adenylate cyclase
are required to elucidate the mechanisms regulating
activity in myocardial tissue was observed. Therefore it
is likely that the increase in FHR after cortisol infusion
In summary, we have demonstrated in fetal sheep
to ADX fetuses results from a stimulatory effect of
that 1) there is a consistent increase in FABP baseline
cortisol directly on the fetal heart. It is also likely that
and a decrease in FHR baseline at 120–140 days GA; 2)
postreceptor events are involved in the changes in FHR
the normal gestational age-dependent increase in FABP
after cortisol infusion. Additionally, the baroreceptors
that occurs in late gestation is attenuated by ADX at
probably play a role in this cortisol-induced increase in
110 days GA in fetal sheep; 3) cortisol infusion begin-
FHR because FHR in ADXϩF fetuses began to increase
ning at 117 days GA to adrenalectomized fetal sheep
on the second day of cortisol infusion, contrasting with
produces a sustained increase in FABP and FHR, which
the FABP increase that occurred immediately on the
is maintained up to 6 days; 4) 24-h rhythms in FABP
first day (Fig. 4, C and F). Thus the early rise in FABP
and FHR exist from 120 to 140 days GA, and their peak
may dampen the mechanisms that lead to the increase
times and amplitudes do not change throughout the
in FHR. In the long-term study, we observed a steady
study period; and 5) the 24-h rhythm in FHR remained
decrease in baseline FHR from 120 to 140 days GA and
unaffected by fetal ADX with or without subsequent
an increase between 140 and 143 days GA (Fig. 1). It is
cortisol supplementation. Taken together, these find-
possible that, in physiological conditions in sheep partu-
ings obtained in the present study indicate that gluco-
rition, the chronotropic effect of cortisol is not strong
corticoids of fetal adrenal origin play an important role
enough to override mechanisms that cause a baseline
in regulating ontogenic changes in baseline FABP
during late gestation in the sheep fetus, whereas their
16. Figueroa, J. P., S. Mahan, E. R. Poore, and P. W. Nathanielsz.
role in baseline FHR regulation does not appear to be
Characteristics and analysis of uterine electromyographic activ-ity in the pregnant sheep. Am. J. Obstet. Gynecol. 151: 524–531,
prominent until 140 days GA, and that the 24-h
rhythms in FHR that exist during the last 3 wk of
17. Graham, D. M., L. D. Longo, and C. Y. Cheung. Catechol-
gestation are not regulated by the fetal adrenal.
amine secretion from the adrenal medulla of the fetus: regulationby hormones. J. Dev. Physiol. (Eynsham) 8: 227–236, 1986.
The authors thank Dr. Norio Shinozuka for data transfer and
18. Hanson, M. A. The control of heart rate and blood pressure in
analysis and Karen Moore for assistance in preparing this paper.
the fetus: theoretical considerations. In: Fetus and Neonate,
This study was supported by National Institute of Child Health
Physiology and Clinical Applications. Cambridge, UK: Cam-
and Human Development Grants HD-28014 and HD-21350.
bridge University Press, 1995, vol. 2, p. 1–22.
Address for reprint requests: P. W. Nathanielsz, Laboratory for
19. Itskovitz, J., E. F. LaGamma, and A. M. Rudolph. Baroreflex
Pregnancy and Newborn Research, Dept. of Physiology, College of
control of the circulation in chronically instrumented fetal lambs.
Veterinary Medicine, Cornell Univ., Ithaca, NY 14853-6401. Circ. Res. 52: 589–596, 1983.
20. Janssen, B. J., C. M. Tyssen, H. Duindam, and W. J.
Received 2 March 1998; accepted in final form 29 September 1998. Rietveld. Suprachiasmatic lesions eliminate 24-h blood pres- sure variability in rats. Physiol. Behav. 55: 307–311, 1994. REFERENCES
21. Janssen, B. J., C. M. Tyssen, and H. A. Struyker-Boudier.
Modification of circadian blood pressure and heart rate variabil-
1. Arduini, D., G. Rizzo, E. Parlati, C. Giorlandino, H. Valen-
ity by five different antihypertensive agents in spontaneously
sise, S. Dell’Acqua, and C. Romanini. Modifications of ultra-
hypertensive rats. J. Cardiovasc. Pharmacol. 17: 494–503, 1991.
dian and circadian rhythms of fetal heart rate after fetal-
22. Kitanaka, T., J. Alonso, R. D. Gilbert, B. L. Siu, G. K.
maternal adrenal gland suppression: a double blind study. Clemons, and L. D. Longo. Fetal responses to long-term Prenat. Diagn. 6: 409–417, 1986.
hypoxemia in sheep. Am. J. Physiol. 256 (Regulatory Integrative
2. Barker, D. J. P., A. R. Bull, C. Osmond, and S. J. Simmonds. Comp. Physiol. 25): R1348–R1354, 1989.
Fetal and placental size and risk of hypertension in adult life. Br.
23. Klein, D. C., R. Y. Moore, and S. M. Reppert. Suprachias- Med. J. 301: 259–262, 1990. matic Nucleus: The Mind’s Clock. New York: Oxford University
3. Benediktsson, R., R. S. Lindsay, J. Noble, J. R. Seckl, and C. R. Edwards. Glucocorticoid exposure in utero: new model for
24. Lawler, F. H., and R. A. Brace. Fetal and maternal arterial
adult hypertension. Lancet 341: 339–341, 1993.
pressures and heart rates: histograms, correlations, and rhythms.
4. Bernardi, M., F. Trevisani, R. De Palma, A. Ligabue, F. Am. J. Physiol. 243 (Regulatory Integrative Comp. Physiol. 12):
Capani, M. Baraldini, and G. Gasbarrini. Chronobiological
evaluation of sympathoadrenergic function in cirrhosis. Relation-
25. Magyar, D. M., F. Fridshal, C. W. Elsner, T. Glaz, J. Eliot,
ship with arterial pressure and heart rate. Gastroenterology 93:
A. H. Klein, K. C. Lowe, J. E. Buster, and P. W. Nathanielsz.
Time-trend analysis of plasma cortisol concentration in the fetal
5. Blanco, C. E., G. S. Dawes, M. A. Hanson, and H. B.
sheep in relation to parturition. Endocrinology 107: 155–159,
McCooke. Carotid baroreceptors in fetal and newborn sheep. Pediatr. Res. 24: 342–346, 1988.
26. Mancia, G., A. Ferrari, L. Gregorini, G. Parati, G. Pomi-
6. Boddy, K., G. S. Dawes, R. Fisher, S. Pinter, and J. S. dorssi, G. Bertinieri, G. Grassi, M. di Rienzo, A. Pedott, Robinson. Fetal respiratory movements, electrocortical and and A. Zanchetti. Blood pressure and heart rate variabilities in
cardiovascular responses to hypoxaemia and hypercapnia in
normotensive and hypertensive human beings. Circ. Res. 53:
sheep. J. Physiol. (Lond.) 243: 599–618, 1974.
7. Brace, R. A., and T. R. Moore. Diurnal rhythms in fetal urine
27. Matthews, J. N. S., D. G. Altman, M. J. Campbell, and P.
flow, vascular pressures, and heart rate in sheep. Am. J. Physiol. Royston. Analysis of serial measurements in medical research.
261 (Regulatory Integrative Comp. Physiol. 30): R1015–R1021,
Br. Med. J. 300: 230–235, 1990.
28. McDonald, T. M., and P. W. Nathanielsz. Bilateral destruction
8. Chan, M. Y., S. Dai, J. H. He, and C. W. Ogle. In-vivo and
of the fetal paraventricular nuclei prolongs gestation in sheep.
in-vitro studies on the effects of chronic dexamethasone treat-
Am. J. Obstet. Gynecol. 165: 764–770, 1991.
ment on cardiovascular responses to sympathetic stimulation.
29. Moura, M. J. C. S., and S. De Moraes. Forced swim stress: Arch. Int. Physiol. Biochim. Biophys. 99: 323–330, 1991.
supersensitivity of the isolated rat pacemaker to the chrono-
9. Dalton, K. J., G. S. Dawes, and J. E. Patrick. Diurnal,
tropic effect of isoprenaline and the role of corticosterone. Gen.
respiratory, and other rhythms of fetal heart rate in lambs. Pharmacol. 25: 1341–1347, 1994. Am. J. Obstet. Gynecol. 127: 414–424, 1977.
30. Myers, D. A., X. Y. Ding, and P. W. Nathanielsz. Effect of fetal
10. Dawes, G. S. The control of fetal heart rate and its variability in
adrenalectomy on messenger RNA for proopiomelanocortin in
lambs. In: Fetal Heart Rate Monitoring, edited by W. Kunzel.
the anterior pituitary and for corticotropin- releasing hormone in
Berlin: Springer-Verlag, 1985, p. 184–190.
the paraventricular nucleus of the ovine fetus. Endocrinology
11. Derks, J. B., D. A. Giussani, S. L. Jenkins, R. A. Wentworth, G. H. Visser, J. F. Padbury, and P. W. Nathanielsz. A
31. Nathanielsz, P. W., A. Bailey, E. R. Poore, G. D. Thorburn,
comparative study of cardiovascular, endocrine and behavioural
and R. Harding. The relationship between myometrial activity
effects of betamethasone and dexamethasone administration to
and sleep state and breathing in fetal sheep throughout the last
fetal sheep. J. Physiol. (Lond.) 499: 217–226, 1997.
third of gestation. Am. J. Obstet. Gynecol. 138: 653–659, 1980.
12. Dodic, M., and E. M. Wintour. Effects of prolonged (48 h)
32. Navarro, H. A., E. M. Kudlacz, and T. A. Slotkin. Control of
infusion of cortisol on blood pressure, renal function and fetal
adenylate cyclase activity in developing rat heart and liver:
fluids in the immature ovine foetus. Clin. Exp. Pharmacol.
effects of prenatal exposure to terbutaline or dexamethasone. Physiol. 21: 971–980, 1994. Biol. Neonate 60: 127–136, 1991.
13. Economides, D. L., K. H. Nicolaides, and S. Campbell.
33. Nelson, W., Y. L. Tong, J. K. Lee, and F. Halberg. Methods for
Metabolic and endocrine finding in appropriate and small for
cosinor rhythmometry. Chronobiologia 6: 305–323, 1979.
gestational age fetuses. J. Perinat. Med. 19: 97–105, 1991.
34. Norman, L. J., and J. R. G. Challis. Synergism between
14. Eijzendach, V., J. H. Sneek, and C. Borst. Arterial pressure
systemic corticotropin-releasing factor and arginine vasopressin
and heart period in the conscious rabbit: diurnal rhythm and
on adrenocorticotropin release in vivo varies as a function of
influence of activity. Clin. Exp. Pharmacol. Physiol. 13: 585–592,
gestational age. Endocrinology 120: 1052–1058, 1987.
35. Ray, N. D., C. S. Turner, N. M. Rawashdeh, and J. C. Rose.
15. Engel, B. T., and M. I. Talan. Diurnal pattern of hemodynamic
Ovine fetal adrenal gland and cardiovascular function. Am. J.
performance in nonhuman primates. Am. J. Physiol. 253 (Regula-Physiol. 254 (Regulatory Integrative Comp. Physiol. 23): R706–
tory Integrative Comp. Physiol. 22): R779–R785, 1987.
36. Robinson, S. R., C. H. Wong, S. S. Robertson, P. W.
hypertension in sheep. Can. J. Physiol. Pharmacol. 65: 1739–
Nathanielsz, and W. P. Smotherman. Behavioral responses of
the chronically instrumented sheep fetus to chemosensory stimuli
44. Stein, H. M., K. Oyama, A. Martinez, B. A. Chappell, E.
presented in utero. Behav. Neurosci. 109: 551–562, 1995. Buhl, L. Blount, and J. F. Padbury. Effects of corticosteroids
37. Sano, H., H. Hayashi, M. Makino, H. Takezawa, M. Hirai, H.
in preterm sheep on adaptation and sympathoadrenal mecha-
Saito, and S. Ebihara. Effects of suprachiasmatic lesions on
nisms at birth. Am. J. Physiol. 264 (Endocrinol. Metab. 27):
circadian rhythms of blood pressure, heart rate and locomotor
activity in the rat. Jpn. Circ. J. 59: 565–573, 1995.
45. Tangalakis, K., E. R. Lumbers, K. M. Moritz, M. K. Tow-
38. Sato, K., F. Chatani, and S. Sato. Circadian and short-term stoless, and E. M. Wintour. Effect of cortisol on blood pressure
variabilities in blood pressure and heart rate measured by
and vascular reactivity in the ovine fetus. Exp. Physiol. 77:
telemetry in rabbits and rats. J. Auton. Nerv. Syst. 54: 236–246,
46. Unno, N., D. A. Giussani, W. K. H. Man A Hing, X. Y. Ding,
39. Schnell, C. R., and J. M. Wood. Measurement of blood pressure J. H. Collins, and P. W. Nathanielsz. Changes in ACTH and cortisol responsiveness following repeated partial umbilical cord
and heart rate by telemetry in conscious unrestrained marmo-
occlusion in the late gestation ovine fetus. Endocrinology 138:
sets. Lab. Anim. Care 29: 258–261, 1995.
40. Scoggins, B. A., J. P. Coghlan, D. A. Denton, W. F. Graham,
47. Wakatsuki, A., Y. Murata, Y. Ninomiya, N. Masaoka, J. G. T. J. Humphery, and J. A. Whitworth. Haemodynamics of Tyner, and K. K. Kutty. Autonomic nervous system regulation
ACTH-induced hypertension in sheep. Clin. Sci. (Colch.) 57:
of baseline heart rate in the fetal lamb. Am. J. Obstet. Gynecol.
41. Simonetta, G., I. R. Young, C. L. Coulter, N. J. Hey, and I. C.
48. Walker, A. M., J. Cannata, M. H. Dowling, B. Ritchie, and McMillan. Fetal adrenalectomy does not affect circulating en- J. E. Maloney. Sympathetic and parasympathetic control of
kephalins in the sheep fetus during late gestation. Neuroendocri-
heart rate in unanaesthetized fetal and newborn lambs. Biol.
42. Smith, T. L., T. G. Coleman, K. A. Stanek, and W. R. Murphy.
49. Wallace, M. J., S. B. Hooper, and R. Harding. Role of the
Hemodynamic monitoring for 24 h in unanesthetized rats.
adrenal glands in the maturation of lung liquid secretory mecha-
Am. J. Physiol. 253 (Heart Circ. Physiol. 22): H1335–H1341,
nisms in fetal sheep. Am. J. Physiol. 270 (Regulatory IntegrativeComp. Physiol. 39): R33–R40, 1996.
43. Spence, C. D., J. P. Coghlan, D. A. Denton, E. H. Mills, M. A.
50. Wood, C. E., C. Y. Cheung, and R. A. Brace. Fetal heart rate, Nelson, J. A. Whitworth, and B. A. Scoggins. Role of the
arterial blood pressure, and blood volume responses to cortisol
autonomic nervous system, renin-angiotensin system, and argi-
infusion. Am. J. Physiol. 253 (Regulatory Integrative Comp.
nine vasopressin during the onset and maintenance of ACTH
Physiol. 22): R904–R909, 1987.
FLORAL AND FAUNAL COMPENDIA FLORAL AND FAUNAL COMPENDIA Species observed at the San Pedro Waterfront Project site during site surveys conducted by Jones & Stokes during October and November 2005 are identified as follows. Pinaceae - Pine Family Aizoaceae - Fig-Marigold Family Amaranthaceae - Amaranth Family Anacardiaceae - Sumac Family Lemonadeberry Apiaceae -
This chapter is not intended to be a ªGuide for Authorsº such as those thatyou can find in any journal. Our main advice is: do not write the paperfirst in your own language and then translate it into English; instead, do itin English directly. When you have a subject that you want to report, first of all you needto look up references. You can refer to the Index Medicus (http://www.ncbi.nlm.nih.
 Blood pressure and heart rate in the ovine fetus:ontogenic changes and effects of fetal adrenalectomy
NOBUYA UNNO,1,2 CHI H. WONG,1,3 SUSAN L. JENKINS,1 RICHARD A. WENTWORTH,1XIU-YING DING,1 CUN LI,1 STEVEN S. ROBERTSON,4 WILLIAM P. SMOTHERMAN,3AND PETER W. NATHANIELSZ11Laboratory for Pregnancy and Newborn Research, Department of Physiology, College of VeterinaryMedicine, and 4Department of Human Development, Cornell University,Ithaca, New York 14853-6401; 3Laboratory of Perinatal Neuroethology, Department of Psychology,Binghamton University, Binghamton, New York 13902; and 2Department of Obstetricsand Gynecology, Faculty of Medicine, University of Tokyo, 113 Tokyo, Japan
Unno, Nobuya, Chi H. Wong, Susan L. Jenkins, Rich-
Blood pressure and heart rate in the ovine fetus:ontogenic changes and effects of fetal adrenalectomy
NOBUYA UNNO,1,2 CHI H. WONG,1,3 SUSAN L. JENKINS,1 RICHARD A. WENTWORTH,1XIU-YING DING,1 CUN LI,1 STEVEN S. ROBERTSON,4 WILLIAM P. SMOTHERMAN,3AND PETER W. NATHANIELSZ11Laboratory for Pregnancy and Newborn Research, Department of Physiology, College of VeterinaryMedicine, and 4Department of Human Development, Cornell University,Ithaca, New York 14853-6401; 3Laboratory of Perinatal Neuroethology, Department of Psychology,Binghamton University, Binghamton, New York 13902; and 2Department of Obstetricsand Gynecology, Faculty of Medicine, University of Tokyo, 113 Tokyo, Japan
Unno, Nobuya, Chi H. Wong, Susan L. Jenkins, Rich-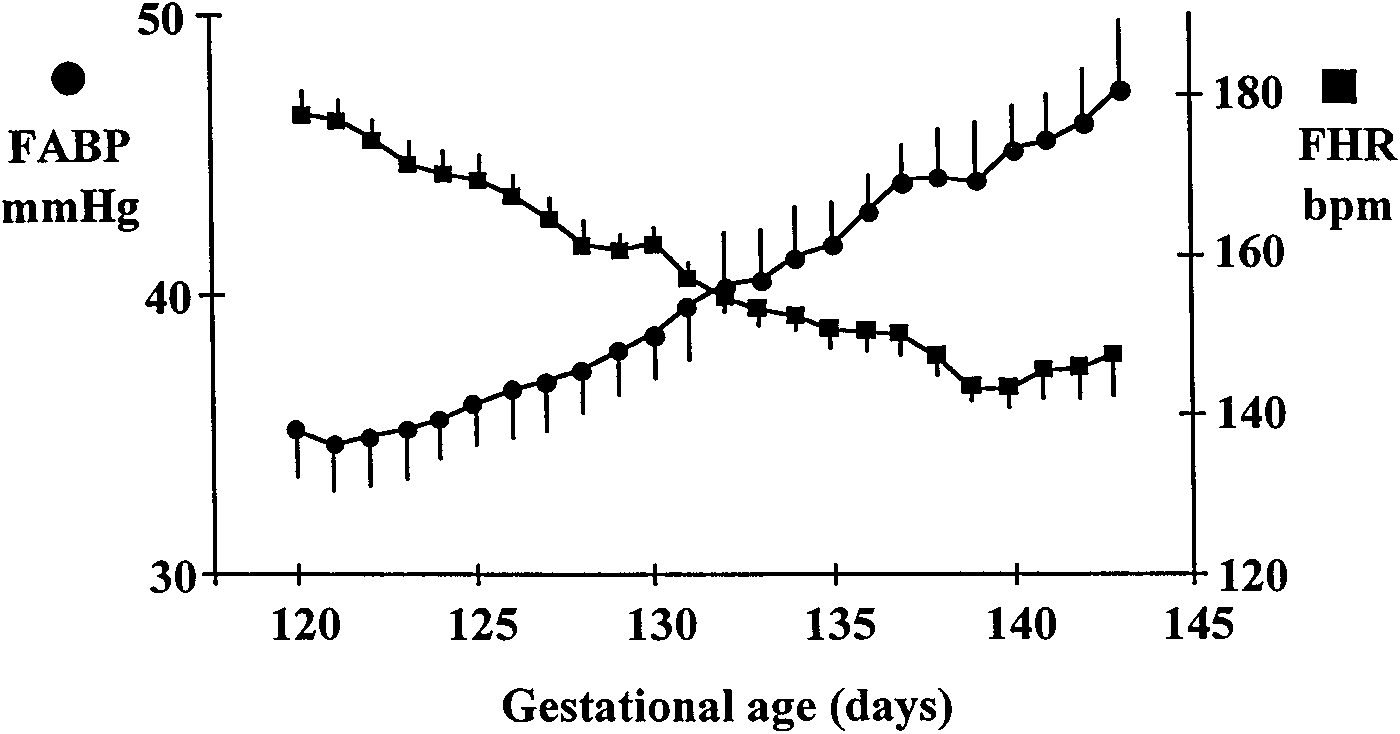
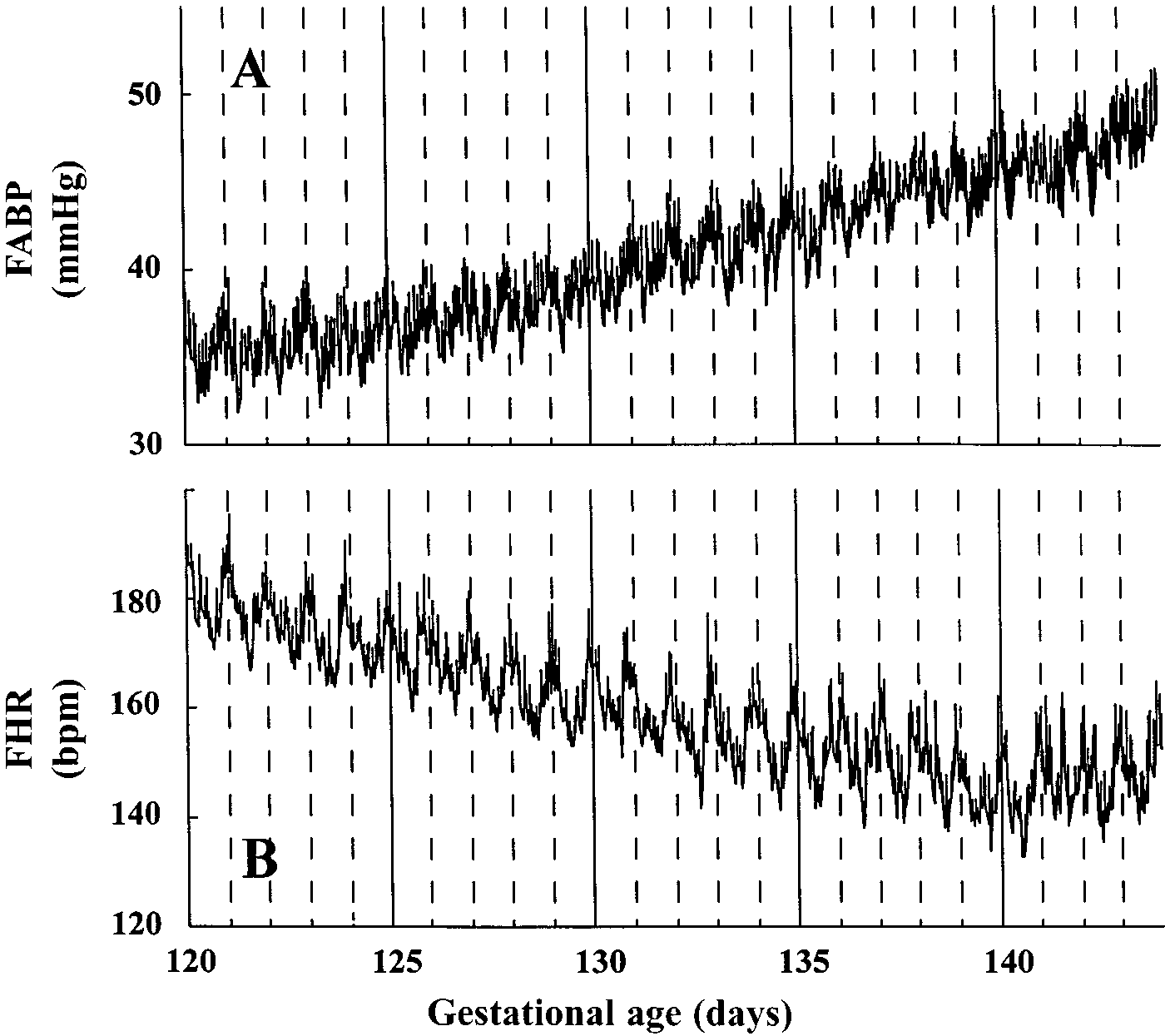 Table 2. Peak time amplitude of 24-h rhythms in FABPand FHR in the long-term study
Values are means Ϯ SE of 5 fetuses, except in fetal arterial blood
pressure (FABP) at 120–126 days GA and 127–133 days GA, wheren ϭ 4. FHR, fetal heart rate. No significant differences were observedthroughout the study period. Lights were on at 0700 and off at 2100.
Table 2. Peak time amplitude of 24-h rhythms in FABPand FHR in the long-term study
Values are means Ϯ SE of 5 fetuses, except in fetal arterial blood
pressure (FABP) at 120–126 days GA and 127–133 days GA, wheren ϭ 4. FHR, fetal heart rate. No significant differences were observedthroughout the study period. Lights were on at 0700 and off at 2100.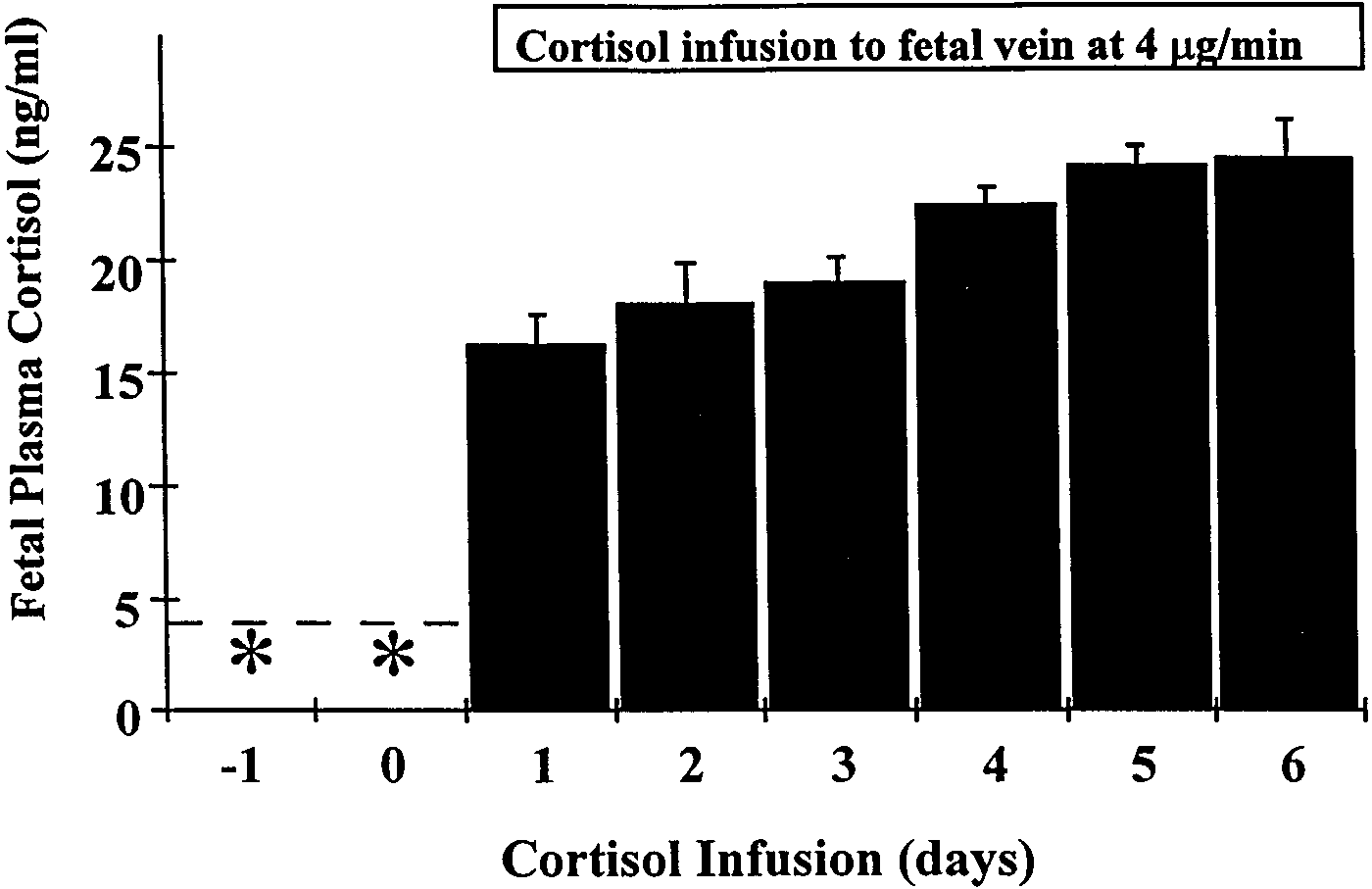 Table 4. Arterial blood gases and pH for fetuses of Con,
Baselines of FABP and FHR. Daily baseline FABP
ADX, and ADX ϩ F in the ADX study
and FHR values on the sixth day after surgery were39.6 Ϯ 1.4 and 190 Ϯ 2, 39.4 Ϯ 1.6 and 185 Ϯ 1, and
44.9 Ϯ 1.9 mmHg and 187 Ϯ 3 beats/min in Con, ADX,
and ADXϩF fetuses, respectively. There were no differ-
ences among these groups. Hourly FABP and FHR
values are shown for Con, ADX, and ADXϩF fetuses in
Changes in FABP and FHR in Con and ADX. During
the study period there was a significant increase in
FABP associated with a significant decrease in FHR in
Con fetuses. In contrast there was no significant change
in FABP in ADX fetuses, whereas the decrease in FHR
was similar to those in Con fetuses.
Table 4. Arterial blood gases and pH for fetuses of Con,
Baselines of FABP and FHR. Daily baseline FABP
ADX, and ADX ϩ F in the ADX study
and FHR values on the sixth day after surgery were39.6 Ϯ 1.4 and 190 Ϯ 2, 39.4 Ϯ 1.6 and 185 Ϯ 1, and
44.9 Ϯ 1.9 mmHg and 187 Ϯ 3 beats/min in Con, ADX,
and ADXϩF fetuses, respectively. There were no differ-
ences among these groups. Hourly FABP and FHR
values are shown for Con, ADX, and ADXϩF fetuses in
Changes in FABP and FHR in Con and ADX. During
the study period there was a significant increase in
FABP associated with a significant decrease in FHR in
Con fetuses. In contrast there was no significant change
in FABP in ADX fetuses, whereas the decrease in FHR
was similar to those in Con fetuses.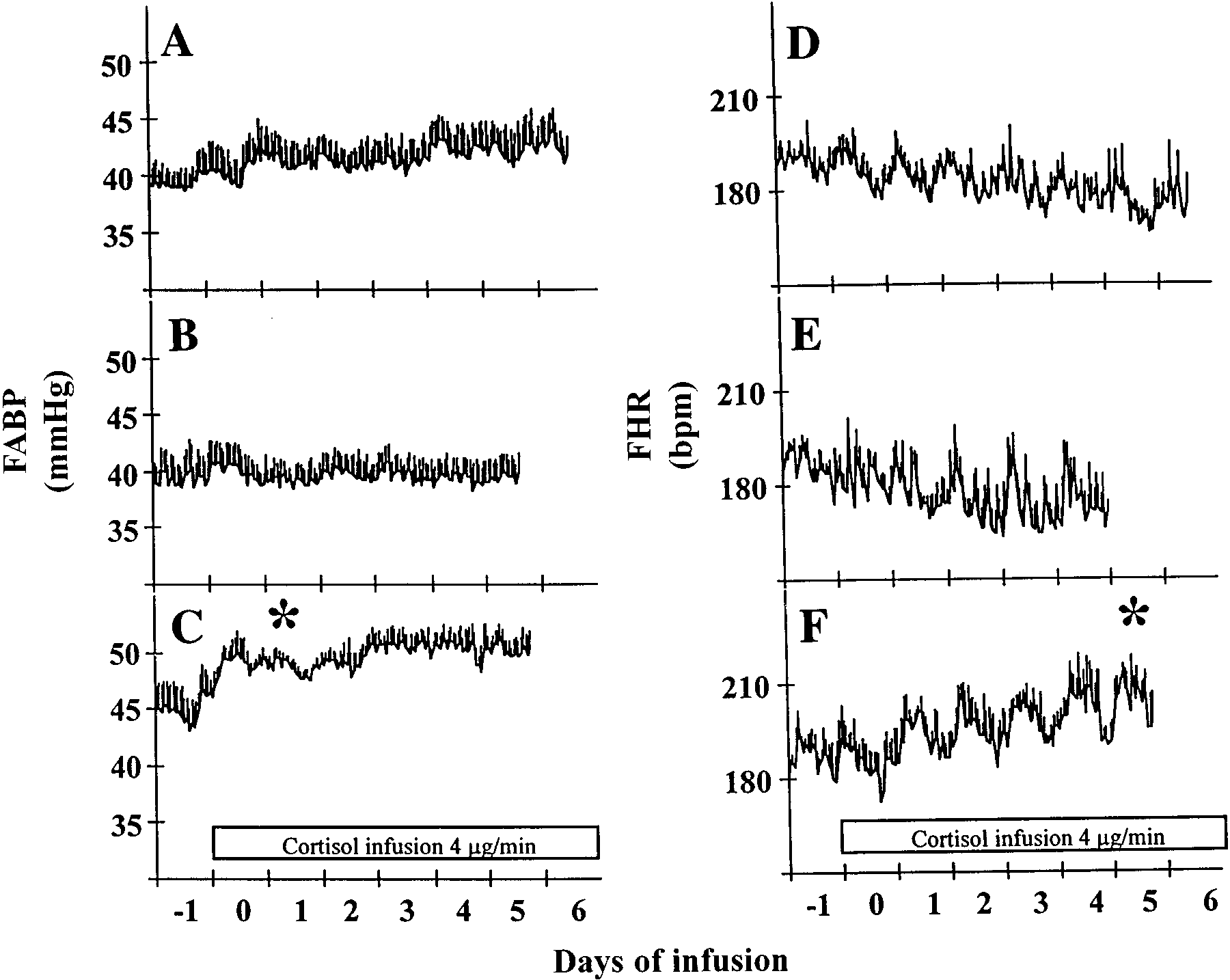 Fig. 4. Hourly FABP and hourly FHR. Values aremeans Ϯ SE beginning on the sixth day (i.e., 24-hbefore the beginning of cortisol and/or vehicleinfusion) in control (Con), ADX, and ADXϩFfetuses. Each day starts at 1600. A: hourly FABPin Con fetuses (n ϭ 5). B: hourly FABP in ADXfetuses (n ϭ 4). C: hourly FABP in ADXϩFfetuses (n ϭ 4); D: hourly FHR in Con fetuses(n ϭ 4); E: hourly FHR in ADX fetuses (n ϭ 3); F:hourly FHR in ADXϩF fetuses (n ϭ 3). Barindicates period of cortisol infusion in C and F.
Fig. 4. Hourly FABP and hourly FHR. Values aremeans Ϯ SE beginning on the sixth day (i.e., 24-hbefore the beginning of cortisol and/or vehicleinfusion) in control (Con), ADX, and ADXϩFfetuses. Each day starts at 1600. A: hourly FABPin Con fetuses (n ϭ 5). B: hourly FABP in ADXfetuses (n ϭ 4). C: hourly FABP in ADXϩFfetuses (n ϭ 4); D: hourly FHR in Con fetuses(n ϭ 4); E: hourly FHR in ADX fetuses (n ϭ 3); F:hourly FHR in ADXϩF fetuses (n ϭ 3). Barindicates period of cortisol infusion in C and F. 36. Robinson, S. R., C. H. Wong, S. S. Robertson, P. W.
36. Robinson, S. R., C. H. Wong, S. S. Robertson, P. W.