Increased expression of cd40 on bone marrow cd34+ hematopoietic progenitor cells in patients with systemic lupus erythematosus: contribution to fas-mediated apoptosis
Vol. 60, No. 2, February 2009, pp 543–552
Increased Expression of CD40 on Bone Marrow CD34ϩ
Hematopoietic Progenitor Cells in Patients With
Katerina Pyrovolaki, Irene Mavroudi, Prodromos Sidiropoulos, Aristides G. Eliopoulos,
Dimitrios T. Boumpas, and Helen A. Papadaki
Objective. Patients with systemic lupus erythem- portion of apoptotic cells and decreased the proportion atosus (SLE) display increased apoptosis of bone mar- of colony-forming cells as compared with untreated row (BM) CD34؉ hematopoietic progenitor cells. This cultures. The CD40L-mediated effects were amplified study was undertaken to evaluate the expression of following treatment with recombinant Fas ligand, sug- CD40 and CD40L in the BM of SLE patients, and to gesting that the effects of these ligands are synergistic. explore the possible involvement of these molecules in CD40L levels were significantly increased in long-term apoptosis of CD34؉ cells. BM culture supernatants and adherent layers of BM Methods. The proportion and survival character- cells from SLE patients as compared with controls. istics of CD40؉ cells within the BM CD34؉ fraction Conclusion. These data reveal a novel role for the from SLE patients and healthy controls were evaluated CD40/CD40L dyad in SLE by demonstrating that up- by flow cytometry. The production of CD40L by BM regulation and induction of CD40 on BM CD34؉ cells stromal cells was assessed using long-term BM cultures, from patients with SLE contribute to the amplification and the effect of CD40L on the survival characteristics of Fas-mediated apoptosis of progenitor cells. and clonogenic potential of CD34؉ cells was evaluated ex vivo by flow cytometry and clonogenic assays.
The emerging role of hematopoietic and mesen-
Results. SLE patients displayed an increased
chymal stem cells in the pathophysiology and treatment
proportion of CD40؉ cells within the CD34؉ fraction
of autoimmune diseases has stimulated interest in the
as compared with controls. The CD34؉CD40؉ sub-
biologic properties of bone marrow (BM)–derived stem
population contained an increased proportion of apo-
cells in these patients (1–4). In this context, we have
ptotic cells compared with the CD34؉CD40؊ fraction
previously shown that patients with systemic lupus ery-
in patients and controls, suggesting that CD40 is in-
thematosus (SLE) display low numbers of BM CD34ϩ
volved in the apoptosis of CD34؉ cells. Stimulation of
hematopoietic progenitor cells, due to induction of
patients’ CD34؉ cells with CD40L increased the pro-
apoptosis by autoreactive T lymphocytes (5). Thesefindings have been further substantiated by studies
Dr. Eliopoulos’ work was supported in part by a grant from
demonstrating low numbers of peripheral blood (PB)
the Association of Cancer Research, UK.
CD34ϩ cells in association with increased apoptosis (6),
Katerina Pyrovolaki, MD, Irene Mavroudi, BSc, Prodromos
Sidiropoulos, MD, Aristides G. Eliopoulos, PhD, Dimitrios T.
as well as abnormal immunophenotypic characteristics
Boumpas, MD, FACP, Helen A. Papadaki, MD, PhD: University of
of BM CD34ϩ cells (7) in SLE patients. In addition,
Crete School of Medicine, Heraklion, Crete, Greece.
clinical data have shown a low PB CD34ϩ progenitor
Address correspondence and reprint requests to Helen A.
Papadaki, MD, PhD, Professor of Hematology, University Hospital of
cell yield in SLE patients undergoing autologous stem
Heraklion, PO Box 1352, Heraklion, Crete, Greece. E-mail: epapadak
cell transplantation, in comparison with that in patients
Submitted for publication May 2, 2008; accepted in revised
Regarding the underlying pathophysiologic mecha-
nisms, it has been shown that the Fas and Fas ligand
in this study. All patients satisfied the American College of
(FasL) systems, belonging to the tumor necrosis factor
Rheumatology revised criteria for the diagnosis of SLE andhad evidence of active disease according to the established
(TNF) receptor (TNFR) and TNF families, respectively,
criteria (27). The SLE Disease Activity Index (SLEDAI) score
is implicated, at least in part, in the apoptotic process of
(scale 0–100) (28) was Ն4 in all patients. The characteristics of
hematopoietic progenitor cells in SLE (5,7). Specifically,
the patients are summarized in Table 1. Patients had discon-
it has been suggested that increased local production of
tinued any medication for at least 24 hours prior to BM
interferon-␥ and FasL by autoreactive T lymphocytes
aspiration. As controls, 20 hematologically healthy subjects,age- and sex-matched with the patients, were studied. Institu-
results in up-regulation of Fas and apoptosis of CD34ϩ
tional ethics committee approval was granted prior to the
study. Informed consent, in accordance with the Declaration of
CD40, a TNFR family member, and its ligand,
Helsinki, was obtained from all patients.
CD40L (CD154) (10), have also been implicated in the
BM sample preparation. BM aspirates (10 ml) ob-
pathophysiology of SLE (11,12). It has been shown that
tained from the posterior iliac crest of patients and healthycontrols were immediately diluted 1:1 in Iscove’s modified
overexpression of CD40L by activated T cells triggers a
Dulbecco’s medium (IMDM; Gibco Invitrogen, Paisley, Scot-
cascade of events in CD40ϩ target cells, resulting in
land) supplemented with 100 IU/ml penicillin–streptomycin
humoral immune dysregulation and autoantibody pro-
(Gibco Invitrogen) and 10 IU/ml preservative-free heparin
duction in SLE (12–15). Interestingly, a biologically
(Sigma-Aldrich, St. Louis, MO). Diluted BM samples werecentrifuged on Histopaque 1077 (density 1.077 gm/cm3; Sigma-
active, membrane-cleaved, soluble form of CD40L
Aldrich) at 400g for 30 minutes at room temperature to obtain
(sCD40L) has been identified and found to be increased
BM mononuclear cells (BMMCs). Cell numbers and viability
in patients’ sera, which contributes to the pathophysiol-
were assessed after staining with trypan blue.
ogy of the disease by further augmenting the CD40-
Immunophenotyping and 7-aminoactinomycin D (7- AAD) staining. An indirect immunofluorescence technique was used to evaluate the expression of CD40 and Fas antigens
The possible distribution of CD40 and its func-
within the BM CD34ϩ cell fraction. Briefly, 1 ϫ 106 BMMCs
tion on hematopoietic progenitor cells in SLE are,
were stained with phycoerythrin (PE)–conjugated mouse
however, entirely unknown. It is of interest that although
anti-human CD34 monoclonal antibody (mAb) (QBEND-10;
CD40–CD40L interactions have been associated with
Immunotech, Marseilles, France) and fluorescein isothiocya-
the rapid expansion of normal B lymphocytes, several
nate (FITC)–conjugated mouse anti-human CD40 (5C3; BDBiosciences-PharMingen, San Jose, CA) or Fas (CD95)
studies have shown that the activation of CD40 in
(LOB 3/17; Serotec, Kidlington, UK) mAb. PE- and FITC-
lymphoma, melanoma, carcinoma cells, and hepatocytes
conjugated mouse IgG isotype-matched controls were used as
results in suppression of cell proliferation and/or induc-
negative controls. Following 30 minutes of incubation on ice,
tion of apoptosis (19–23). One of the mechanisms
cells were washed twice in phosphate buffered saline
proposed to explain the proapoptotic function of CD40
(PBS)/1% fetal calf serum (FCS) (Gibco Invitrogen)/0.05%azide and were further stained with 7-AAD (Calbiochem-
in malignant cells and hepatocytes is the CD40-inducible
Novabiochem, La Jolla, CA) for the evaluation of the propor-
up-regulation of Fas (23–26). However, whether such a
tion of apoptotic cells, as previously described (29). Briefly,
cooperative interaction between the CD40 and Fas
100 l 7-AAD solution (200 g/ml) was added to the cells,
pathways is also implicated in the apoptosis of hemato-
followed by suspension in 1 ml PBS and incubation on ice for
poietic stem cells in SLE is unknown. Furthermore,
20 minutes in the dark. Following centrifugation, the superna-tant was removed, and cells were fixed in 500 l 2% parafor-
although several studies have demonstrated increased
maldehyde solution (Sigma-Aldrich). Unstained fixed cells
amounts of circulating sCD40L in SLE, the cytokine
levels in the SLE BM microenvironment and the possi-
Cell samples were analyzed on an Epics Elite model
ble pathophysiologic significance of these cytokines in
flow cytometer (Coulter, Miami, FL) within 30 minutes of
SLE have so far not been investigated. Therefore, the
fixation. Data were acquired and processed on 500,000 eventsto evaluate the proportion of CD34ϩ cells within the BMMCs,
present study was performed to evaluate the expression
the percentage of CD40ϩ and Fasϩ cells in the CD34ϩ cell
of CD40/CD40L in BM hematopoietic progenitor cells
fraction, and the proportion of 7-AAD–negative (live),
and the BM microenvironment in patients with SLE, and
7-AAD–dim (apoptotic), and 7-AAD–bright (dead) cells
to explore the possible involvement of this receptor/
within the CD40- or Fas-expressing CD34ϩ cells.
ligand dyad in the apoptotic depletion of CD34ϩ cells. Effect of CD40L on the survival characteristics and clonogenic potential of CD34؉ cells. Flow cytometric analysis. CD34ϩ cells were isolated from the BMMCs of SLE patients PATIENTS AND METHODS
and healthy controls by magnetic-activated cell sorting(MACS) analysis (MACS isolation kit; Miltenyi Biotec, Ber-
Patients. Twenty patients with SLE (19 women and 1
gisch Gladbach, Germany) according to the manufacturer’s
man; median age 43 years [range 19–73 years]) were enrolled
protocol. The purity of the CD34ϩ cells was more than 96% in
EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
Clinical and laboratory characteristics of the patients with systemic lupus erythematosus*
* SLEDAI ϭ Systemic Lupus Erythematosus Disease Activity Index (scale 0–100); ANA ϭ antinuclear antibody; anti-dsDNA ϭ antibodies againstdouble-stranded DNA; Hgb ϭ hemoglobin; CNS ϭ central nervous system; PDN ϭ prednisolone; HCQ ϭ hydroxychloroquine; CYC ϭcyclophosphamide; MMF ϭ mycophenolate mofetil; ND ϭ not determined.
all experiments. We then cultured CD34ϩ cells in 96 round-
results were expressed as the total number of colony-forming
bottomed well plates at a density of 1 ϫ 104 CD34ϩ cells/well
in the presence of 1 g/ml recombinant human CD40L
Long-term BM cultures (LTBMCs). For preparation
(rHuCD40L) (BMS308/2; Bender MedSystems, Vienna, Aus-
of LTBMCs, 107 BMMCs were grown according to a standard
tria) and/or 1 g/ml rHuFasL (BMS309/2; Bender MedSys-
technique (30), in 10 ml IMDM supplemented with 10% FCS,
tems) in 200 l IMDM/1% FCS. Following 48 hours of
10% horse serum (Gibco Invitrogen), 100 IU/ml penicillin–
incubation, the cells were stained with 7-AAD in the same
streptomycin, 2 mM L-glutamine, and 10Ϫ6M hydrocortisone
manner as described above, for the evaluation of the propor-
sodium succinate (Sigma-Aldrich), and incubated at 33°C in a
tion of live, apoptotic, and dead CD34ϩ cells. In a number of
fully humidified atmosphere of 5% CO . At weekly intervals,
experiments, Fas antigen expression was also evaluated on
cultures were examined for stromal layer formation, using an
CD34ϩ cells that had been treated in the same manner as
inverted microscope, and were fed by removing half of the
described above, with 1 g/ml rHuCD40L.
medium and replacing it with an equal volume of fresh IMDM
Clonogenic assay. We cultured 3 ϫ 103 CD34ϩ cells
supplemented as described above. At weeks 3–4, when a
from SLE patients and normal controls in 1 ml IMDM
confluent stromal layer was formed, cell-free supernatants
supplemented with 30% FCS, 1% bovine serum albumin
were stored at Ϫ72°C for the determination of sCD40L, using
(BSA) (Gibco Invitrogen), 10Ϫ4M mercaptoethanol (Sigma-
a commercially available (Bender MedSystems) enzyme-linked
Aldrich), 0.075% sodium bicarbonate (Gibco Invitrogen),
immunoabsorbent assay (ELISA). The sensitivity of the assay
2 mM L-glutamine (Sigma-Aldrich), and 0.9% methylcellulose
is 0.6 ng/ml. The culture samples were tested by ELISA after
(StemCell Technologies, Vancouver, British Columbia, Can-
being concentrated using a Speedvac method (31).
ada), in the presence of 5 ng granulocyte–macrophage colony-
For the assessment of CD40L expression in LTBMCs
stimulating factor (R&D Systems, Minneapolis, MN), 50 ng
of stroma, total messenger RNA (mRNA) was extracted from
interleukin-3 (R&D Systems), and 2 IU erythropoietin
the adherent cells of confluent LTBMCs, using the RNeasy
(Janssen-Cilag, Bucks, UK) in the presence or absence of 1 g
mini kit (Qiagen, Hilden, Germany) according to the manu-
rHuCD40L and/or 1 g rHuFasL. Cultures were set up in
facturer’s instructions. Contaminating DNA was removed by
duplicate in 35-mm petri dishes and incubated at 37°C in a fully
digestion with RNAse-free DNAse. The SuperScript Pream-
humidified atmosphere of 5% CO . In a separate set of
plification System (Gibco Invitrogen) was used for first-strand
experiments, 105 BMMCs were cultured as described above, in
complementary DNA (cDNA) synthesis from 1 g total RNA,
the presence or absence of 1 g/ml CD40L neutralizing
followed by reverse transcription–polymerase chain reaction
antibody (MK13A4; Bender MedSystems) or with the same
(PCR) with specific primers. PCR products were normalized
quantity of PBS as a control. In all cases, on day 14 of the
according to the amount of  -microglobulin ( m) in the
cultures, colonies were scored as previously described, and
samples. We performed one tube reaction, and the primer
Flow cytometric analysis of BMMCs in SLE patients and healthy controls*
CD34ϩFasϩ cell fraction, 7-AADdim cells
CD34ϩFasϪ cell fraction, 7-AADdim cells
CD34ϩCD40ϩ cell fraction, 7-AADdim cells
CD34ϩCD40Ϫ cell fraction, 7-AADdim cells
* Except where indicated otherwise, values are the mean Ϯ SD. BMMC ϭ bone marrow mononuclear cell; SLE ϭ systemic lupus erythematosus;7-AADdim ϭ 7-aminoactinomycin D–dim (apoptotic).
concentration of CD40L was 7 times higher than the concen-
Table 2. In accordance with previously reported data (5),
the proportion of CD34ϩ cells within the BMMC frac-
The forward and reverse primers were 5Ј-AGAATC-
CTCAAATTGCGGC-3Ј and 5Ј-TGTGGGTATTTGCAGCT-
tion was statistically significantly lower in SLE patients
CTG-3Ј, respectively, for CD40L (PCR product size 286 bp),
(mean Ϯ SD 1.14 Ϯ 0.35%) compared with healthy
and 5Ј-TCCAACATCAACATCTTGGT-3Ј and 5Ј-TCCCCC-
controls (2.06 Ϯ 0.62%; P Ͻ 0.0001), and the proportion
AAATTCTAAGCAGA-3Ј, respectively, for  m (PCR prod-
of Fasϩ cells within the CD34ϩ cell compartment was
uct size 123 bp). Conditions for 37 cycles of PCR amplificationfollowing initial denaturation were 94°C for 30 seconds, 62°C
significantly higher in patients (20.69 Ϯ 19.53%) com-
for 30 seconds, and 72°C for 45 seconds. PCR products were
pared with controls (7.16 Ϯ 4.46%; P ϭ 0.0138). The
electrophoresed on a 1.5% agarose gel and visualized under
percentage of CD40-expressing CD34ϩ cells was signif-
ultraviolet light by ethidium bromide staining. The positive
icantly increased in SLE patients (10.79 Ϯ 6.90%)
control was cDNA from the Ms5-CD40L–transfected cell line(provided by one of us [AGE]). Results were analyzed using
compared with healthy controls (5.41 Ϯ 2.89%; P ϭ
the ImageJ analysis program (National Institutes of Health,
0.0061). Moreover, a highly significant correlation was
found between the proportion of CD40-expressing
Statistical analysis. Data were analyzed using the
CD34ϩ cells and the proportion of Fas-expressing
GraphPad Prism statistical program (GraphPad Software, SanDiego, CA) by means of the nonparametric Mann-Whitney
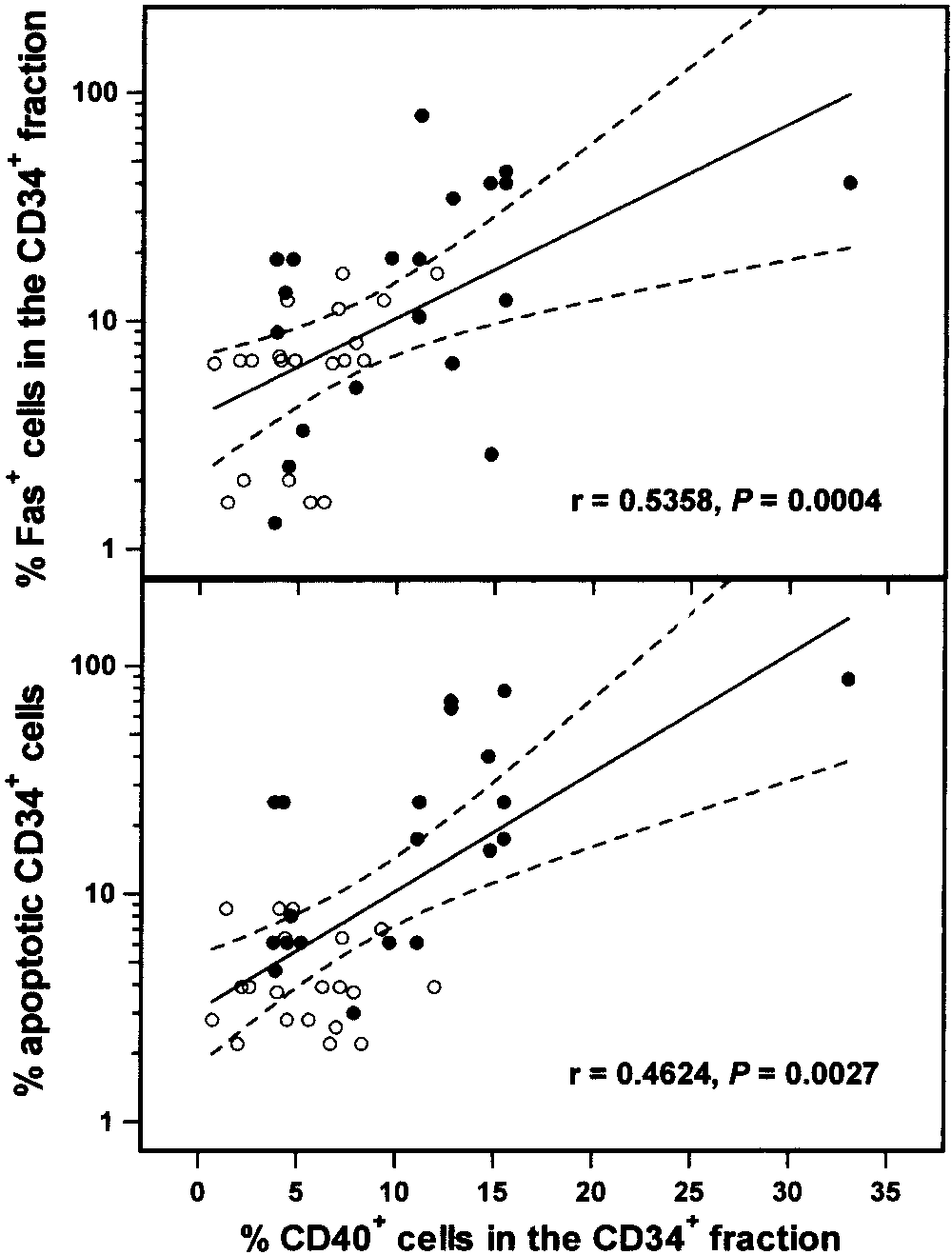
CD34ϩ cells (r ϭ 0.5358, P ϭ 0.0004), suggesting a
and Spearman’s tests. Student’s t-test for paired samples was
parallel mode of expression of the 2 molecules in the
used to compare differences in the proportion of Fasϩ cells
study cell population (Figure 1). Notably, no statistically
and the number of CFCs between cultures treated and those
significant difference was found between patients and
not treated with rHuCD40L or CD40L neutralizing antibody. Group data are expressed as the mean Ϯ SD.
controls in the proportion of CD40ϩ cells detected inthe non-CD34ϩ BMMC fraction (7.03 Ϯ 2.65% and6.13 Ϯ 2.43%, respectively; P ϭ 0.818). Survival characteristics of the CD34؉CD40؉ Expression of CD40 and Fas antigens on CD34؉ cells. Data on the survival characteristics of the patients’ cells. Results from flow cytometric analysis of the
CD34ϩ cells are presented in Table 2. In accordance
CD34ϩ cells from patients with SLE are presented in
with the results from previously reported studies (5), the
EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
prominent role of the Fas antigen in the apoptosis ofBM CD34ϩ cells (5,32,33). Interestingly, an increasedrate of apoptosis was also observed in the CD34ϩFasϪcell fraction in the patients compared with the controls(P ϭ 0.0066), suggesting that additional mechanismsother than up-regulation of Fas antigen expression areoperating in the apoptotic depletion of CD34ϩ cells inSLE.
To investigate the possible involvement of CD40
in the apoptotic process of CD34ϩ cells, we studied thesurvival characteristics of the CD40ϩ and CD40Ϫ cellsubpopulations. In the gate of CD34ϩ cells, the propor-tion of apoptotic cells was significantly higher among theCD40ϩ cells than among the CD40Ϫ cells, in both thepatients (37.43 Ϯ 18.24% versus 11.85 Ϯ 9.72%; P Ͻ0.0001) and the controls (23.07 Ϯ 10.03% versus 5.41 Ϯ6.41%; P Ͻ 0.0001) (Table 2). In the gate of CD34Ϫcells, however, no statistically significant difference inthe percentage of apoptotic cells was documented be-tween the CD40ϩ and CD40Ϫ cell subpopulations, ineither the patients (2.40 Ϯ 2.42% versus 3.30 Ϯ 3.65%;
P ϭ 0.5792) or the controls (2.47 Ϯ 1.80% versus 1.65 Ϯ1.64%; P ϭ 0.638). Furthermore, among all of thesubjects studied, a highly significant correlation wasnoted between the proportion of CD40ϩ cells and thepercentage of apoptotic cells within the CD34ϩ sub-population (r ϭ 0.4624, P ϭ 0.0027) (Figure 1), but not
Figure 1. Correlations between the proportion of CD40ϩ cells and
within the non-CD34ϩ subpopulation (r ϭ 0.1887, P ϭ
the percentages of Fasϩ cells (top) and apoptotic cells (bottom) within
0.818). These results indicate that overexpression of the
the CD34ϩ cell fraction of bone marrow from patients with systemic
CD40 antigen may have a role in the apoptotic depletion
lupus erythematosus (n ϭ 20; solid circles) and healthy subjects (n ϭ20; open circles), as determined by linear regression analysis. Regres-
of BM CD34ϩ cells in patients with SLE.
sion lines and the 95% confidence intervals are shown as solid lines
All of the patients studied had active disease.
However, CD40 expression on CD34ϩ cells did notcorrelate with parameters of disease activity, such as theSLEDAI score, albumin levels, antinuclear antibody
proportion of apoptotic cells within the CD34ϩ com-
titers, and C3 values. Nevertheless, there was a correla-
partment was significantly increased in SLE patients
tion with hemoglobin levels (r ϭ Ϫ0.6689, P ϭ 0.0013),
(26.03 Ϯ 26.40%) compared with healthy controls
further indicating a possible apoptosis-inducing role of
(4.47 Ϯ 2.21%; P Ͻ 0.0001). The percentage of apopto-
CD40 on hematopoiesis in patients with SLE.
tic cells was higher in the CD34ϩFasϩ cell compart-
Effect of CD40L on CD34؉ hematopoietic pro-
ment compared with the CD34ϩFasϪ cell compart-
genitor cell death. To investigate the hypothesis that the
ment, in both the SLE patients (43.50 Ϯ 24.56% versus
CD40 antigen may directly and/or indirectly have a role
22.89 Ϯ 28.38%; P ϭ 0.0098) and the healthy controls
in the apoptotic process of CD34ϩ cells, we evaluated
(25.60 Ϯ 14.15% versus 3.24 Ϯ 2.25%; P Ͻ 0.0001),
the proportion of apoptotic cells following incubation of
whereas no significant difference was documented in the
purified CD34ϩ cells from SLE patients (n ϭ 3) and
proportion of apoptotic cells detected in the Fasϩ and
healthy controls (n ϭ 3) with rHuCD40L and/or rHu-
FasϪ subpopulations of the non-CD34ϩ cell compart-
FasL. The results are presented in Figure 2A. In SLE
ment, in either the patients (2.87 Ϯ 2.60% and 3.74 Ϯ
patients, the addition of rHuFasL significantly increased
3.97%, respectively; P ϭ 0.6848) or the controls (1.89 Ϯ
the proportion of apoptotic cells (33.93 Ϯ 3.52%) com-
2.07% and 1.62 Ϯ 1.35%, respectively; P ϭ 0.7452).
pared with untreated (baseline) cultures (24.27 Ϯ
These findings further highlight the previously reported
3.78%; P ϭ 0.0317). The addition of rHuCD40L also
apoptotic cells compared with that in untreated cultures(53.26 Ϯ 11.23%; P ϭ 0.0013), compared with that incultures treated with rHuFasL alone (P ϭ 0.0466), orcompared with that in cultures treated with rHuCD40Lalone (P ϭ 0.0024). In healthy controls, the addition ofrHuCD40L and/or rHuFasL did not result in statisticallysignificant increases in the proportion of apoptotic cellsas compared with that in untreated cultures, probablybecause of the low levels of expression of surface CD40and Fas on normal CD34ϩ cells.
These data suggest that CD40L displays an indi-
rect, rather than a direct, apoptosis-inducing effect onCD40-expressing CD34ϩ cells, by facilitating the FasL-mediated apoptotic process. To substantiate this hy-pothesis, we evaluated Fas antigen expression on immu-nomagnetically sorted CD34ϩ cells from SLE patients(n ϭ 12) following a 48-hour incubation with rHuCD40L. We found that the proportion of Fasϩ cells significantlyincreased in cultures treated with rHuCD40L (48.60 Ϯ12.63%) as compared with that in untreated cultures(23.71 Ϯ 7.47%; P Ͻ 0.0001). The up-regulation of Fasexpression by CD40L may therefore explain, at least inpart, the amplifying effects of CD40L on Fas-mediatedCD34ϩ cell death. Effect of CD40L on the clonogenic potential of CD34؉ cells. To further characterize the effect of CD40L on the function of CD34ϩ cells, we investigated the clonogenic potential of CD34ϩ cells from SLE patients (n ϭ 11) and healthy controls (n ϭ 11) in the presence of rHuCD40L and/or rHuFasL. Results are depicted in Figure 2B.
In cultures of patients’ cells treated with rHu-
FasL, a significant decrease in CFC number (36.54 Ϯ
Figure 2. Effect of CD40L on the survival characteristics and clono-
25.46 CFCs per 3 ϫ 103 CD34ϩ cells) was observed in
genic potential of CD34ϩ cells. A, Proportion of apoptotic CD34ϩ cells in the bone marrow of patients with systemic lupus erythematosus
comparison with that in untreated cultures (62.09 Ϯ
(SLE) (n ϭ 3) and healthy subjects (n ϭ 3) following 48-hour
22.09 CFCs per 3 ϫ 103 CD34ϩ cells; P ϭ 0.0418). In
incubation of the cells with recombinant human Fas ligand (rhFasL) (1
the presence of rHuCD40L, CFC numbers were simi-
g/ml) and/or recombinant human CD40L (1 g/ml). Apoptosis was
larly decreased (45.73 Ϯ 28.16 CFCs per 3 ϫ 103 CD34ϩ
assessed by flow cytometry and 7-aminoactinomycin D staining. B,
cells) as compared with baseline; however, the differ-
Numbers of colony-forming cells (CFCs) per 3 ϫ 103 CD34ϩ cells,determined using a clonogenic assay, in patients with SLE (n
ence was not statistically significant. A profound de-
healthy subjects (n ϭ 11) following 14-day culture of the cells with
crease in CFC number was observed in the presence of
recombinant human FasL (1 g/ml) and/or recombinant human CD40L
both rHuCD40L and rHuFasL (18.45 Ϯ 12.10 CFCs per
(1 g/ml). In both series of experiments, treated cultures were compared
3 ϫ 103 CD34ϩ cells) as compared with that in un-
with untreated cultures (baseline) using the nonparametric Mann-
treated cultures (P ϭ 0.0001) or compared with that in
Whitney test. Bars show the mean and SD.
cultures treated with rHuCD40L alone (P ϭ 0.0256) orrHuFasL alone (P ϭ 0. 0459). Clonogenic assays using
increased the proportion of apoptotic cells (30.20 Ϯ
CD34ϩ cells from healthy subjects demonstrated that
1.47%), but not to a statistically significant extent as
rHuCD40L and/or rHuFasL reduced the number of
compared with untreated cultures (P ϭ 0.0645). The
CFCs as compared with that in untreated cultures.
addition of both rHuCD40L and rHuFasL resulted in a
However, these differences were not statistically signif-
statistically significant increase in the proportion of
icant. Collectively, these data suggest that CD40L exerts,
EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
ng/ml) as compared with that in healthy controls (0.27 Ϯ0.18 ng/ml; P ϭ 0.0061). Increase in clonogenic potential of SLE BM progenitor cells by neutralization of CD40L. To further substantiate the involvement of the CD40 pathway in BM progenitor cell survival in SLE patients, we evalu- ated the clonogenic potential of patient and normal BMMCs in the presence or absence of a CD40L neu- tralizing antibody. On the basis of our observations indicating that CD40 was overexpressed on the patients’ CD34ϩ cells in the BMMC fraction (Table 2) and that CD40L was overproduced by cells in the BM micro- Figure 3. Concentration of CD40L in long-term bone marrow cul-
environment (Figure 3), it was anticipated that neutral-
tures (LTBMCs). A, Cells from SLE patients (n ϭ 20) and healthy
ization of CD40L would increase the CFC number selec-
controls (n ϭ 20) were harvested at confluence, and levels of CD40L,upon concentration in LTBMC supernatants, were measured by
tively in SLE patient samples, but not in the controls.
enzyme-linked immunosorbent assay. The mean concentration and
95% confidence intervals are indicated by horizontal lines and shaded areas, respectively. B, CD40L mRNA expression was determined in LTBMC adherent layers of cells from all SLE patients and healthy controls. Bars show the mean and SD cumulative data. Groups were compared using the nonparametric Mann-Whitney test. See Figure 2 for other definitions.
mainly, a synergistic effect on the FasL-induced impair-ment of the clonogenic potential of CD40-expressingCD34ϩ cells, rather than a direct negative effect. Increased CD40L expression in SLE LTBMCs.
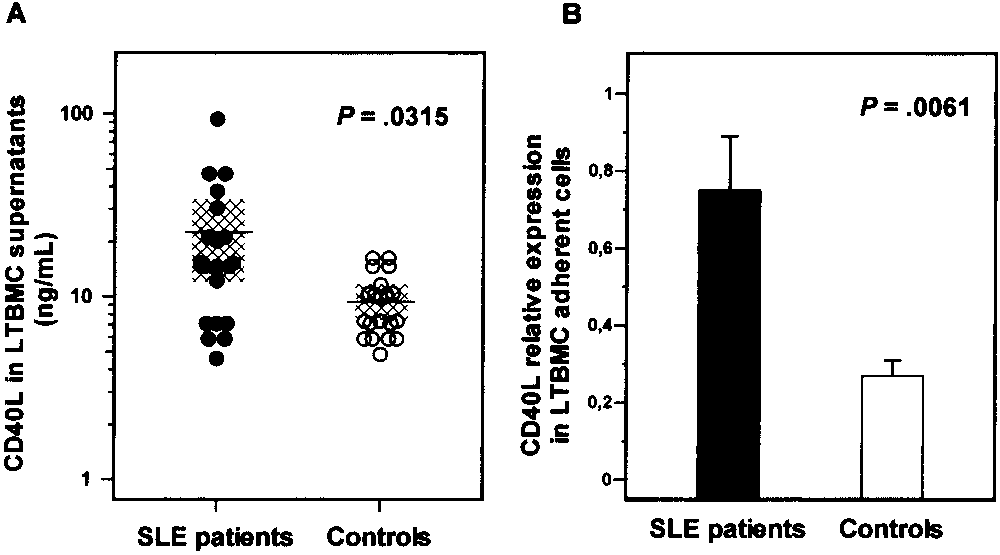
To probe the pathophysiologic significance of the in-creased expression of CD40 on CD34ϩ cells frompatients with SLE, we evaluated CD40L levels andCD40L mRNA expression in LTBMCs, an experimentaldesign that represents an in vitro model mimicking theBM microenvironment (30). The levels of sCD40L werefirst determined in LTBMC supernatants. Results arepresented in Figure 3A.
SLE patients displayed increased sCD40L levels
(21.97 Ϯ 21.07 ng/ml) as compared with healthy controls(9.42 Ϯ 3.63 ng/ml; P ϭ 0.0315), suggesting that localproduction of sCD40L is increased in the BM of SLEpatients. Individual sCD40L levels inversely correlated
Figure 4. Effects of neutralization of CD40L on the clonogenic po-
with the proportion of CD34ϩ cells in the subjects
tential of bone marrow progenitor cells from patients with SLE. Bone
studied (r ϭ Ϫ0.6139, P Ͻ 0.0001), further indicating the
marrow mononuclear cells (BMMCs) (105) from SLE patients (n ϭ 6)
negative effect of sCD40L on the reserves of BM
and healthy controls (n ϭ 6) were cultured using a standard clonogenicassay, in 1 ml Iscove’s modified Dulbecco’s medium supple-
mented with 30% fetal calf serum, 1% bovine serum albumin, 10Ϫ4M
To further substantiate the increased local pro-
mercaptoethanol, 0.075% sodium bicarbonate, 2 mM L-glutamine, 0.9%
duction of CD40L in SLE BM, we evaluated the cyto-
methylcellulose, 5 ng granulocyte–macrophage colony-stimulating
kine mRNA expression in LTBMC adherent cell ex-
factor, 50 ng interleukin-3, and 2 IU erythropoietin in the presence
tracts from patients and healthy controls. Cumulative
or absence of 1 g CD40L neutralizing antibody. Bars show the meanand SD number of CFCs obtained in SLE patients and controls in
data are shown in Figure 3B. Consistent with the ELISA
antibody-treated and untreated (baseline) cultures. Groups were com-
data shown in Figure 4A, CD40L mRNA expression was
pared using Student’s t-test for paired samples. See Figure 2 for other
significantly increased in SLE patients (0.75 Ϯ 0.62
obtained from SLE patients (n ϭ 6) resulted in a
displayed an increased proportion of apoptotic, Fas-
statistically significant increase in the clonogenic poten-
expressing cells within the CD34ϩ cell compartment as
tial of BMMCs (93 Ϯ 47 CFCs per 105 BMMCs) as
compared with the the levels in healthy subjects.
compared with that in cultures that did not receive
To explore the possible association of CD40
anti-CD40L antibody (70 Ϯ 48 CFCs per 105 BMMCs;
up-regulation with the apoptotic depletion of the hema-
P ϭ 0.005) or that in cultures with the same quantity of
topoietic progenitor cells in SLE patients, we per-
PBS (69 Ϯ 46 CFCs per 105 BMMCs; P ϭ 0.003). In
formed a subset analysis of the survival characteristics of
contrast, neutralization of CD40L had no impact on the
CD34ϩ cells on the basis of CD40 surface expression.
clonogenic potential of BMMCs obtained from control
We found that the proportion of apoptotic CD34ϩ cells
subjects (n ϭ 6) (159 Ϯ 16 CFCs per 105 BMMCs versus
was significantly increased in the CD40ϩ cells as com-
162 Ϯ 17 CFC per 105 BMMCs in untreated cultures;
pared with the CD40Ϫ cell fraction, in both patients and
P ϭ 0.286) (Figure 4). Thus, our findings suggest that
healthy controls, suggesting a possible involvement of
CD40L is functionally expressed in BMMCs.
CD40 in the apoptotic process of BM hematopoieticprogenitor cells. In favor of this hypothesis was thestrong correlation between the proportion of CD40ϩ
DISCUSSION
cells and the percentage of apoptotic CD34ϩ cells in the
There is increasing evidence that immune dys-
study population. In contrast, CD40 antigen expression
regulation in patients with SLE may disturb the mor-
did not affect the survival characteristics of cells in the
phology and function of the BM microenvironment and
CD34Ϫ BMMC fraction, which comprises mainly the
may severely affect BM hematopoiesis (9,34). Auto-
reactive lymphocytes and proinflammatory mediators
CD40 ligation appears to have opposing effects
have been primarily implicated in BM damage, in that
on cell viability, depending on the cell target (11).
these elements suppress the hematopoiesis-supporting
Interestingly, CD40-dependent apoptotic processes have
capacity of BM stromal cells and accelerate the apo-
been reported to be mediated via up-regulation of other
ptosis of hematopoietic stem/progenitor cells (5). A
TNFR family molecules, including Fas (22,24). More-
varying degree of “unexplained” cytopenias, extreme
over, a cooperative interaction between CD40 and Fas in
cases of aplastic anemia, and insufficient CD34ϩ cell
the promotion of the apoptotic process has been dem-
yield upon mobilization are abnormalities attributed to
onstrated in human hepatocytes (23), B lymphoma cells
the immune-mediated damage of BM in SLE (35,36). A
(38), and carcinoma cells (24). Our data demonstrating
better understanding of the mechanisms that induce the
that incubation of patients’ CD34ϩ cells with CD40L
stem/progenitor cell damage in SLE would have major
resulted in up-regulation of surface Fas indicate, for the
clinical implications in SLE, in view of the current
first time, that a cooperative mechanism between the
interest in exploring the use of autologous stem cells in
CD40 and Fas pathways also exists in BM CD34ϩ cells.
the treatment of severely affected patients (37), since
In accordance with previously reported data (5),
elucidating the effects of these cells may explain poten-
SLE patients were found to display an increased propor-
tial problems in the mobilization, harvesting, and mar-
tion of Fasϩ cells in the CD34ϩ BMMC fraction, and
incubation of purified CD34ϩ cells with FasL resulted in
In the present study, we have investigated the
a significant increase in the proportion of apoptotic cells,
involvement of CD40–CD40L interactions in the apo-
suggesting that functional Fas expression takes place in
ptotic process of the BM hematopoietic stem/progenitor
the BM. The strong correlation between the proportion
cells in patients with SLE. Although these 2 TNFR/TNF
of CD40- and Fas-expressing CD34ϩ cells in SLE
family members are aberrantly expressed in patients’ PB
patients indicates a possible cooperative mode of action
lymphocytes and serum, they have never been studied in
of these molecules in accelerating progenitor cell death.
the BM. We found that CD40 was minimally expressed
Evidence to support this hypothesis comes from our
on normal hematopoietic progenitor cells; however, the
observation that ex vivo treatment of patients’ CD34ϩ
proportion of CD40-expressing CD34ϩ cells was signif-
cells with the combination of CD40L and FasL resulted
icantly increased in SLE patients. In accordance with
in amplification of apoptosis as compared with that in
our previously reported findings on the reserves and
cultures exposed to each ligand alone.
survival characteristics of BM CD34ϩ cells in SLE (5),
It is of interest that treatment of patients’ CD34ϩ
we found that SLE patients had a significantly lower
cells with CD40L alone resulted in a low level of apo-
percentage of CD34ϩ cells in the BMMC fraction and
ptosis induction, suggesting that CD40 activation per se
EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
may affect CD34ϩ cell survival. However, a number of
reduce autoantibody production but also to ameliorate
observations suggest that CD40 activation confers pre-
cytopenias associated with BM dysfunction. The intra-
dominantly indirect negative effects on the survival of
cellular pathways and signaling cascades involved in this
BM progenitor cells through the amplification of Fas-
intriguing crosstalk between CD40 and Fas, the possible
induced apoptosis. Thus, exposure of patients’ CD34ϩ
implication of other TNF/TNFR family members, as
cells to rHuCD40L resulted in up-regulation of Fas
well as the effect of the treatment on CD40 expression
expression. Moreover, we have shown that in vitro
are interesting areas for further investigation.
exposure to a combination of CD40L and FasL induceda profound reduction in the number of CFCs in SLEpatients as compared with treatment with each cytokine
ACKNOWLEDGMENTS
alone, and that the relative reduction in CFC numbers
We thank the staff of the Departments of Rheumatol-
was higher following FasL treatment as compared with
ogy (especially Dr. Heraklis Kritikos) and Hematology at the
University Hospital of Heraklion for providing the BM sam-
It has been suggested that elevated levels of
ples from patients and healthy controls.
sCD40L in SLE patient sera, in association with in-creased and prolonged expression of the membrane
AUTHOR CONTRIBUTIONS
form of the molecule on patients’ T cells, may contribute
Dr. Papadaki had full access to all of the data in the study and
to the pathophysiology of the disease by amplifying the
takes responsibility for the integrity of the data and the accuracy of the
aberrant activation signals through the target cell CD40
data analysis. Study design. Eliopoulos, Papadaki.
(16). In our study, we evaluated sCD40L in LTBMC
Acquisition of data. Pyrovolaki, Mavroudi, Sidiropoulos, Boumpas,
supernatants of patients’ cells, reflecting the cytokine
levels within the BM microenvironment. We found that
Analysis and interpretation of data. Eliopoulos, Papadaki.
culture supernatant levels of sCD40L were significantly
Manuscript preparation. Eliopoulos, Boumpas, Papadaki. Statistical analysis. Papadaki.
increased in patients compared with controls, and thatthese levels correlated inversely with the proportion ofCD34ϩ cells. This finding coupled with the observed
REFERENCES
CFC recovery following CD40L neutralization in pa-
1. Ikehara S. Bone marrow transplantation: a new strategy for
tient, but not control, BMMC cultures further highlight
intractable diseases. Drugs Today (Barc) 2002;38:103–11.
the negative effect that the CD40 pathway may exert on
2. Burt RK, Traynor AE, Craig R, Marmont AM. The promise of
hematopoietic stem cell transplantation for autoimmune diseases.
hematopoiesis in patients with SLE. Consistent with the
Bone Marrow Transplant 2003;31:521–4.
elevated sCD40L levels in the supernatants was the
3. Dazzi F, van Laar JM, Cope A, Tyndall A. Cell therapy for
increased cytokine mRNA expression in LTBMC adher-
autoimmune diseases. Arthritis Res Ther 2007;9:206.
4. Corcione A, Benvenuto F, Ferretti E, Giunti D, Cappiello V,
ent layers derived from SLE patients compared with that
Cazzanti F, et al. Human mesenchymal stem cells modulate B-cell
in normal LTBMCs. The cellular origin of CD40L in the
BM microenvironment in SLE is unknown. CD40L might
5. Papadaki HA, Boumpas DT, Gibson FM, Jayne DR, Axford JS,
be produced by a variety of cells that constitute the
Gordon-Smith EC, et al. Increased apoptosis of bone marrowCD34(ϩ) cells and impaired function of bone marrow stromal
LTBMC adherent layers, such as monocyte/macrophages,
cells in patients with systemic lupus erythematosus. Br J Haematol
endothelial cells, and fibroblasts (11), but also might be
produced by lymphocytes that persist in LTBMC condi-
6. Westerweel PE, Luijten RK, Hoefer IE, Koomans HA, Derksen
RH, Verhaar MC. Haematopoietic and endothelial progenitor
cells are deficient in quiescent systemic lupus erythematosus. Ann
In summary, this study is the first to evaluate the
expression of CD40/CD40L in the BM progenitor cell
7. Sun LY, Zhou KX, Feng XB, Zhang HY, Ding XQ, Jin O, et al.
Abnormal surface markers expression on bone marrow CD34(ϩ)
microenvironment in patients with SLE and healthy
cells and correlation with disease activity in patients with systemic
subjects. Our data reveal a novel role for CD40 in BM
lupus erythematosus. Clin Rheumatol 2007;26:2073–9.
homeostasis in SLE, by demonstrating that up-
8. Statkute L, Verda L, Oyama Y, Traynor A, Villa M, Shook T, et
regulation of the CD40 pathway amplifies Fas-mediated
al. Mobilization, harvesting and selection of peripheral blood stemcells in patients with autoimmune diseases undergoing autologous
CD34ϩ hematopoietic progenitor cell death and con-
hematopoietic stem cell transplantation. Bone Marrow Transplant
tributes to defective BM progenitor cell reserve and
function, both of which are characteristic of SLE. These
9. Papadaki HA. Autoreactive T-lymphocytes are implicated in the
pathogenesis of bone marrow failure in patients with systemic
results further substantiate the rationale of CD40/
lupus erythematosus. Leuk Lymphoma 2003;44:1301–7.
CD40L blockade as treatment of SLE, not only to
10. Fiumara P, Younes A. CD40 ligand (CD154) and tumour necrosis
factor-related apoptosis inducing ligand (Apo-2L) in haematologi-
26. Lee JK, Seki N, Sayers TJ, Subleski J, Gruys EM, Murphy WJ, et
cal malignancies. Br J Haematol 2001;113:265–74.
al. Constitutive expression of functional CD40 on mouse renal
11. Schonbeck U, Libby P. The CD40/CD154 receptor/ligand dyad.
cancer cells: induction of Fas and Fas-mediated killing by CD40L.
12. Koshy M, Berger D, Crow MK. Increased expression of CD40
27. Tan EM, Cohen AS, Fries JF, Masi AT, McShane DJ, Rothfield
ligand on systemic lupus erythematosus lymphocytes. J Clin Invest
NF, et al. The 1982 revised criteria for the classification of systemic
lupus erythematosus. Arthritis Rheum 1982;25:1271–7.
13. Desai-Mehta A, Lu L, Ramsey-Goldman R, Datta SK. Hyper-
28. Bombardier C, Gladman DD, Urowitz MB, Caron D, Chang DH,
expression of CD40 ligand by B and T cells in human lupus and
and the Committee on Prognosis Studies in SLE. Derivation of the
its role in pathogenic autoantibody production. J Clin Invest
SLEDAI: a disease activity index for lupus patients. Arthritis
14. Yellin MJ, Thienel U. T cells in the pathogenesis of systemic lupus
erythematosus: potential roles of CD154-CD40 interactions and
29. Philpott NJ, Turner AJ, Scopes J, Westby M, Marsh JC, Gordon-
costimulatory molecules. Curr Rheumatol Rep 2000;2:24–31.
Smith EC, et al. The use of 7-amino actinomycin D in identifying
15. Yazdany J, Davis J. The role of CD40 ligand in systemic lupus
apoptosis: simplicity of use and broad spectrum of application
erythematosus. Lupus 2004;13:377–80.
compared with other techniques. Blood 1996;87:2244–51.
16. Vakkalanka RK, Woo C, Kirou KA, Koshy M, Berger D, Crow
30. Coutinho LH, De Wynter EA, Will A, Testa NG. Clonal and
MK. Elevated levels and functional capacity of soluble CD40
long-term cultures using human bone marrow. In: Testa NG,
ligand in systemic lupus erythematosus sera. Arthritis Rheum
Milineux G, editors. Haemopoiesis: a practical approach. Oxford:
Oxford University Press; 1993. p. 75–106.
17. Kato K, Santana-Sahagun E, Rassenti LZ, Weisman MH, Tamura
31. Toure R, Kheadr E, Lacroix C, Moroni O, Fliss I. Production of
N, Kobayashi S, et al. The soluble CD40 ligand sCD154 in systemic
antibacterial substances by bifidobacterial isolates from infant
lupus erythematosus. J Clin Invest 1999;104:947–55.
stool active against Listeria monocytogenes. J Appl Microbiol
18. Goules A, Tzioufas AG, Manousakis MN, Kirou KA, Crow MK,
Routsias JG. Elevated levels of soluble CD40 ligand (sCD40L) in
32. Nagafuji K, Shibuya T, Harada M, Mizuno S, Takenaka K,
serum of patients with systemic autoimmune diseases. J Auto-
Miyamoto T, et al. Functional expression of Fas antigen (CD95)
on hematopoietic progenitor cells. Blood 1995;8:883–9.
19. Funakoshi S, Longo DL, Beckwith M, Conley DK, Tsarfaty G,
33. Papadaki HA, Kritikos HD, Gemetzi C, Koutala H, Marsh JC,
Tsarfaty I, et al. Inhibition of human B-cell lymphoma growth by
Boumpas DT, et al. Bone marrow progenitor cell reserve and
CD40 stimulation. Blood 1994;83:2787–94.
function and stromal cell function are defective in rheumatoid
20. Baker MP, Eliopoulos AG, Young LS, Armitage RJ, Gregory CD,
arthritis: evidence for a tumor necrosis factor ␣-mediated effect.
Gordon J. Prolonged phenotypic, functional, and molecular
change in group I Burkitt lymphoma cells on short-term exposure
34. Voulgarelis M, Giannouli S, Tasidou A, Anagnostou D, Ziakas
to CD40 ligand. Blood 1998;92:2830–43.
PD, Tzioufas AG. Bone marrow histological findings in systemic
21. Von Leoprechting A, van der Bruggen P, Pahl HL, Aruffo A,
lupus erythematosus with hematologic abnormalities: a clinico-
Simon JC. Stimulation of CD40 on immunogenic human malig-
pathological study. Am J Hematol 2006;81:590–7.
nant melanomas augments their cytotoxic T lymphocyte-mediated
35. Tagoe C, Shah A, Yee H, Belmont HM. Aplastic anemia in
lysis and induces apoptosis. Cancer Res 1999;59:1287–94.
22. Georgopoulos NT, Steele LP, Thomson MJ, Selby PJ, Southgate J,
systemic lupus erythematosus: a distinct presentation of acquired
Trejdosiewicz LK. A novel mechanism of CD40-induced apoptosis
aplastic anemia? J Clin Rheumatol 2001;7:377–83.
of carcinoma cells involving TRAF3 and JNK/AP-1 activation.
36. Burt RK, Fassas A, Snowden J, van Laar JM, Kozak T, Wulffraat
Cell Death Differ 2006;13:1789–801.
NM, et al. Collection of hematopoietic stem cells from patients
23. Afford SC, Randhawa S, Eliopoulos AG, Hubscher SG, Young
with autoimmune diseases. Bone Marrow Transplant 2001;28:
LS, Adams DH. CD40 activation induces apoptosis in cultured
human hepatocytes via induction of cell surface fas ligand expres-
37. Tyndall A, Daikeler T. Autologous hematopoietic stem cell trans-
sion and amplifies fas-mediated hepatocyte death during allograft
plantation for autoimmune diseases. Acta Haematol 2005;114:
rejection. J Exp Med 1999;189:441–6.
24. Eliopoulos AG, Davies C, Knox PG, Gallagher NJ, Afford SC,
38. Xerri L, Bouabdallah R, Devilard E, Hassoun J, Stoppa AM, Birg
Adams DH, et al. CD40 induces apoptosis in carcinoma cells
F. Sensitivity to Fas-mediated apoptosis is null or weak in B-cell
through activation of cytotoxic ligands of the tumor necrosis factor
non-Hodgkin’s lymphomas and is moderately increased by CD40
superfamily. Mol Cell Biol 2000;20:5503–15.
ligation. Br J Cancer 1998;78:225–32.
25. Zhou F, Ajuebor MN, Beck PL, Le T, Hogaboam CM, Swain MG.
39. Papadaki HA, Marsh JC, Eliopoulos GD. Bone marrow stem cells
CD154-CD40 interactions drive hepatocyte apoptosis in murine
and stromal cells in autoimmune cytopenias. Leuk Lymphoma
fulminant hepatitis. Hepatology 2005;42:372–80.
COMMITTEE FOR THE CORPORATE GOVERNANCE OF LISTED COMPANIES CODE OF CONDUCT TRANSLATION (for reference purposes only) Towards the end of 1998, I thought that a series of favourable conditions was existingin Italy for convening a Committee with the aim of drawing up a Report on CorporateGovernance of listed companies and drafting a Code of Conduct. The market capitalisation of th
Goat Parasites: Management and Control Taken from presentations during the ECA Symposium on Goat Health Revised September 2004 by Mary C. Smith DVM A. Epidemiology Goats are very sensitive to the effects of internal parasitism (such as anemia and low blood protein). Parasitism can cause decreased fertility, abortion, unthriftiness, increased susceptibility to disease, and death. Th
 EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
prominent role of the Fas antigen in the apoptosis ofBM CD34ϩ cells (5,32,33). Interestingly, an increasedrate of apoptosis was also observed in the CD34ϩFasϪcell fraction in the patients compared with the controls(P ϭ 0.0066), suggesting that additional mechanismsother than up-regulation of Fas antigen expression areoperating in the apoptotic depletion of CD34ϩ cells inSLE.
EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
prominent role of the Fas antigen in the apoptosis ofBM CD34ϩ cells (5,32,33). Interestingly, an increasedrate of apoptosis was also observed in the CD34ϩFasϪcell fraction in the patients compared with the controls(P ϭ 0.0066), suggesting that additional mechanismsother than up-regulation of Fas antigen expression areoperating in the apoptotic depletion of CD34ϩ cells inSLE.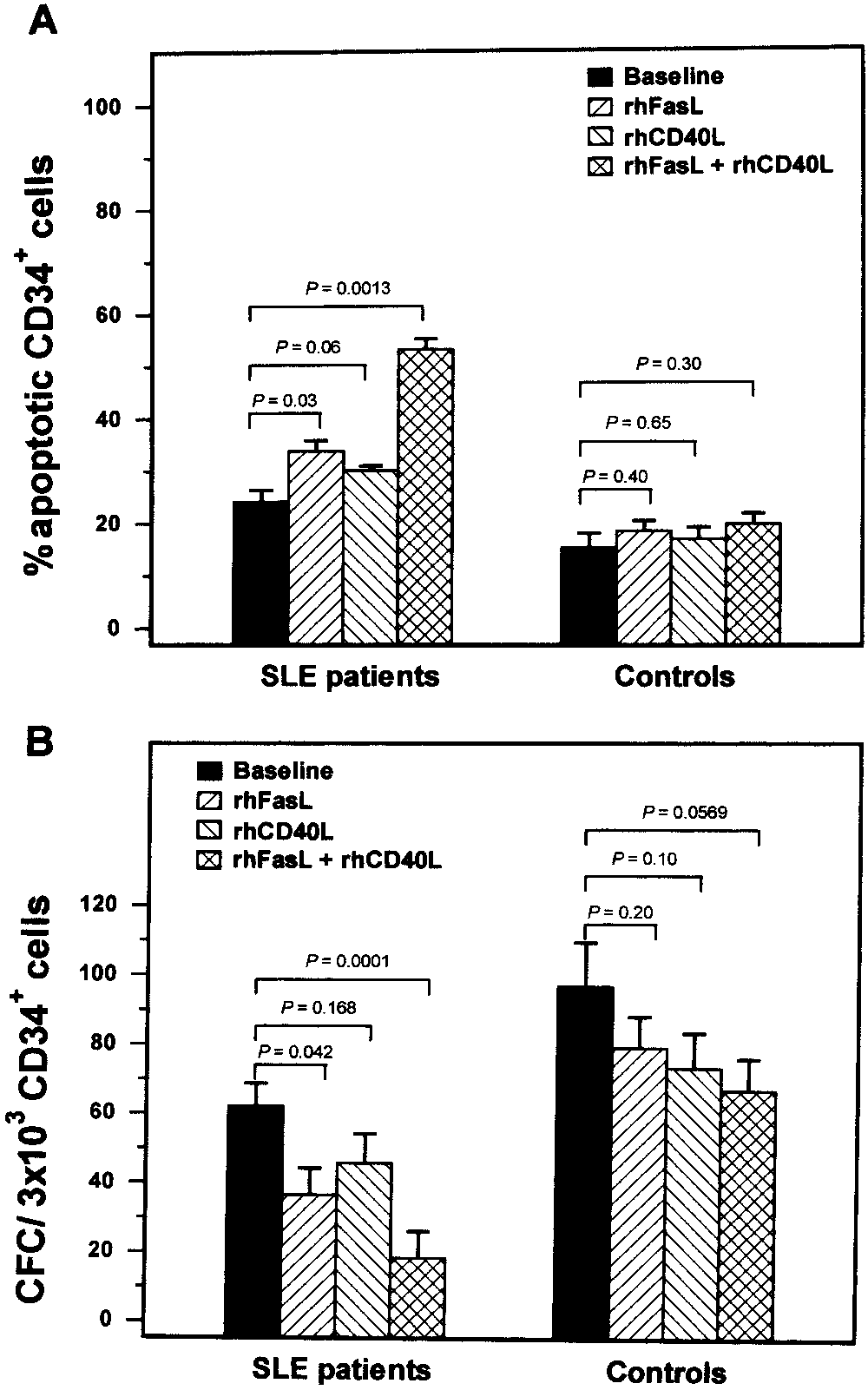 apoptotic cells compared with that in untreated cultures(53.26 Ϯ 11.23%; P ϭ 0.0013), compared with that incultures treated with rHuFasL alone (P ϭ 0.0466), orcompared with that in cultures treated with rHuCD40Lalone (P ϭ 0.0024). In healthy controls, the addition ofrHuCD40L and/or rHuFasL did not result in statisticallysignificant increases in the proportion of apoptotic cellsas compared with that in untreated cultures, probablybecause of the low levels of expression of surface CD40and Fas on normal CD34ϩ cells.
apoptotic cells compared with that in untreated cultures(53.26 Ϯ 11.23%; P ϭ 0.0013), compared with that incultures treated with rHuFasL alone (P ϭ 0.0466), orcompared with that in cultures treated with rHuCD40Lalone (P ϭ 0.0024). In healthy controls, the addition ofrHuCD40L and/or rHuFasL did not result in statisticallysignificant increases in the proportion of apoptotic cellsas compared with that in untreated cultures, probablybecause of the low levels of expression of surface CD40and Fas on normal CD34ϩ cells.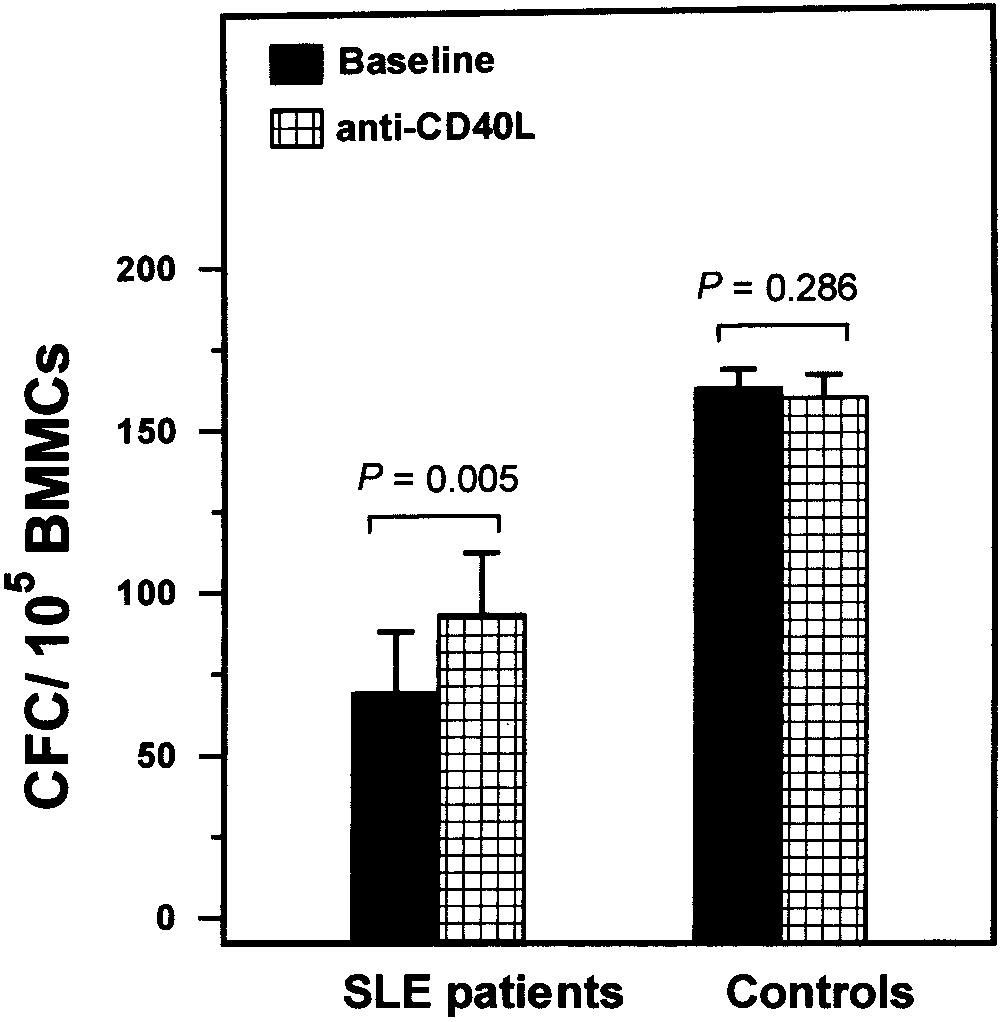
 EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
ng/ml) as compared with that in healthy controls (0.27 Ϯ0.18 ng/ml; P ϭ 0.0061).
EXPRESSION OF CD40 AND Fas-MEDIATED APOPTOSIS OF BM CD34ϩ CELLS IN SLE
ng/ml) as compared with that in healthy controls (0.27 Ϯ0.18 ng/ml; P ϭ 0.0061).