The operational sex ratio of the sea urchin paracentrotus lividus populations: the case of the mediterranean marine protected area of ustica island (tyrrhenian sea, italy)
The operational sex ratio of the sea urchin Paracentrotuslividus populations: the case of the Mediterranean marineprotected area of Ustica Island (Tyrrhenian Sea, Italy)Paola Gianguzza1, Fabio Badalamenti2, Fabrizio Gianguzza3, Chiara Bonaviri1 & Silvano Riggio1
1 Dipartimento di Ecologia, Universita` degli Studi di Palermo, Palermo, Italy2 CNR-IAMC Laboratorio di Biologia Marina, Castellammare del Golfo (Tp), Italy3 Dipartimento di Biologia Cellulare e dello Sviluppo Universita` degli Studi di Palermo, Palermo, Italy
Adult sex ratio; echinoid; Marine ProtectedArea; natural predation; starfish; Tyrrhenian
We investigated, with a series of field and laboratory observations, the possible
effect of the starfish Marthasteria glacialis predation on the operational sexratio (OSR), i.e. the number of sexually mature males divided by the total
number of sexually mature adults of both sexes at any one time, of the edible
Paola Gianguzza, Dipartimento di Ecologia,
sea urchin Paracentrotus lividus. The OSR was estimated three times during the
Universita` degli Studi di Palermo, Via Archirafi
sea urchin summer spawning period (July 2004, June 2005 and July 2006) on
18, 90123 Palermo, Italy. E-mail: [email protected]
barren substrates of Ustica Island Marine Protected Area (Southern TyrrhenianSea, Italy). Four sites were selected: two characterized by high M. glacialis den-
sity (take zone C) and two controls with low starfish density (no-take zone A). Mature sea urchins were independently collected by SCUBA diving and sexed.
The adult sex ratio was skewed towards males at high M. glacialis density sites,whereas it was balanced (1:1) at predator low-density sites. Results of sex-selec-tive feeding experiments in the laboratory showed that females of P. lividuswere more vulnerable to M. glacialis predation. These outcomes underline thepossible importance of M. glacialis in regulating the OSR of P. lividus popula-tions.
tance in determining the structure of rocky reefs
(Verlaque 1987; Sala et al. 1998) and to its economic
The operational sex ratio, hereafter OSR, is defined by
value (Boudouresque & Verlaque 2001). It has been sug-
Emlen & Oring (1977) as the number of sexually mature
gested that its grazing may play a paramount role in the
males divided by the total number of sexually mature and
transition from macroalgal beds to ‘coralline barrens’,
active adults of both sexes at a given moment. It is a
characterized by lower complexity and diversity (Verlaque
major factor influencing the intensity of sexual selection
1987; Sala et al. 1998). At the same time, P. lividus is
(Clutton-Brock & Parker 1992). A skewed OSR usually
intensively exploited in many Mediterranean areas, as
leads to increased competition for mates among members
both male and female gonads are considered a delicacy
of the more abundant sex, while members of the other
(Lawrence 1987; Boudouresque & Verlaque 2001).
sex may have a greater opportunity to exercise mate
In July 2002, during the preliminary survey phase of
a project aimed at defining the role of sea urchin P. lividus
and Arbacia lixula (L.) (Echinodermata: Echinoidea) pop-
(Echinodermata: Echinoidea) has been the object of a
ulations in the process of formation and maintenance of
number of studies in the Mediterranean Sea. The increas-
barrens in the Ustica Island Marine Protected Area
ing interest in this species is primarily due to its impor-
(MPA), we found a male-skewed P. lividus OSR at the
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
take zone C (high starfish M. glacialis density and a
particular in the take area of the MPA (authors’ personal
balanced OSR at the no-take zone A (low starfish den-
observation). Unfortunately, due to the extremely cryptic
sity). Given the ecological and economic importance of
behaviour and nocturnal activity of this species (Savy
P. lividus, this operational sex-ratio disparity was consid-
1987b), abundance estimates are problematic and may be
ered worthy of further investigation. The apparent imbal-
biased. However, a high number of M. glacialis have been
ance in the OSR may reflect a sampling procedure bias,
observed predating sea urchins in recent years, and the
but if this pattern is confirmed, this may have important
remains of sea urchins eaten by starfish are frequently
consequences for fluctuations in population size and
recorded, especially at the barren grounds of zone C
structure, especially in a small population (Emlen &
M. glacialis is a slow-moving predator, able to detect
P. lividus is a species with a chromosomal (genetic) sex
its prey using olfaction (Valentincic 1973). It could be
determination mechanism expected to allocate equal
hypothesized that during the P. lividus spawning period
effort to producing male and female offspring because
the starfish is more attracted by (and hence preferentially
random meiosis would lead to a mean sex ratio of 0.5
consumes) females. The aim of this study was to explore
(Lipani et al. 1996). In this case, environmental condi-
possible variability in OSR of P. lividus populations
tions, such as pronounced temperature fluctuations,
inhabiting barren rocky substrates of Ustica Island MPA.
which may influence sex determination in other echi-
By comparing areas with different densities of M. glacialis,
noids (see Pearse & Cameron 1991), were not involved in
low (the no-take zone A) versus high (take zone C), we
assessed the potential importance of this predator in reg-
According to Clutton-Brock & Vincent (1991), the
ulating the OSR of the P. lividus population of the Ustica
OSR of species with chromosomal sex determination can
Island MPA. Furthermore, with a series of laboratory
be influenced by the following factors: the adult sex ratio
choice experiments we tested the prediction that M. gla-
in the population, the spatial and temporal distribution
cialis prefers to consume P. lividus females rather than
of the two sexes, their differences in growth and mortal-
ity, their potential reproductive output (i.e. number ofsperm delivered and eggs deposited) and reproductivebehaviour. Possible deviation from a 1:1 sex ratio may
also be due to different habitat (i.e. higher food availabil-
ity conditions typical of fringe habitats) that could attracthigher female numbers, as reported before in other
The study was carried out on Ustica Island (MPA) in the
echinoid species (McPherson 1965, 1968; Bernard 1977;
Lessios 1979). Natural selective predation and fishing or
10°43¢43¢¢ E–38°42¢20¢¢ N), 60 km north of the Sicilian
harvesting (in terms of sex and size) also affect the adult
coast (Fig. 1). The MPA, created in 1986, encompasses a
sex ratio and consequently the OSR of marine popula-
total area of 16,000 ha and contains three zones with dif-
ferent degrees of protection. The no-take zone (zone A)
In the Mediterranean Sea, several predators such as
covers 65 ha along the western part of the island, while
fishes, lobsters, starfishes and gastropods are listed amongthe most important sea urchin predators (Savy 1987a),and human harvesting may affect the abundance and sizeof targeted species and have positive indirect conse-quences on algal assemblages (Guidetti & Sala 2007).
Gianguzza et al. (2006) showed that in the MPA of
Ustica Island a regulated human harvesting (50 P. lividusindividuals per person per day, during the summer sea-son) may affect the size and have positive indirect conse-quences on non-target species such as A. lixula. AsP. lividus is a species that lacks sexually dimorphic struc-tures, it is not expected to exhibit differential vulnerabilityto human predation. However, other predators such asthe spiny starfish Marthasteria glacialis (L.) could beinvolved in explaining the observed OSR differences.
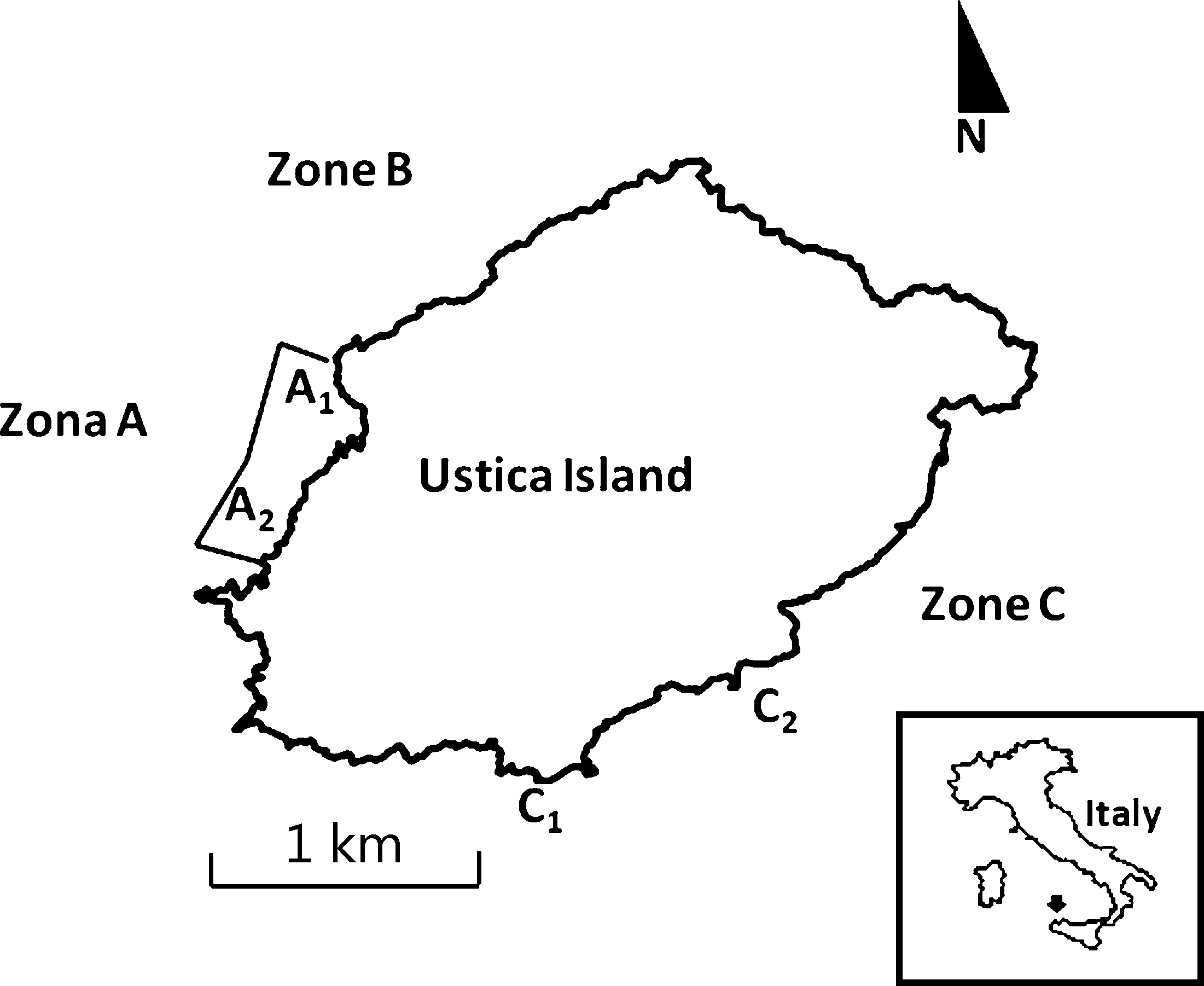
There is evidence that in the last 10 years, the
Fig. 1. Zonation of the Ustica Island MPA and study sites. (A1: Punta
M. glacialis population at Ustica Island has grown, in
Megna; A2: Acquario; C1: Punta dell’Arpa; C2: Punta S. Paolo).
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
the general reserve (zone B) and the take zone (zone C)
seven randomly located quadrats of 1 m2 on shallow
(about 4–5 m depth) rocky barren substrates mostly
According to the institutive decree of the MPA (D.I.
composed of gently sloping platforms. Sampling was con-
12 ⁄ 11 ⁄ 1986), only research activities are authorized in the
ducted on days with good sea-weather conditions, around
no-take zone A, whereas recreational activities, such as
SCUBA diving, boat anchoring, swimming and angling,
We collected sea urchins larger than 40–45 mm (test
are strictly prohibited. There are no restrictions on recrea-
diameter) and thus every year we analysed different
tional activities in either the B or the C zones. Further-
mature animals belonging to this size class. Collected ani-
more, the law authorizes local professional fishing in
mals were separated in numbered landing nets and
these zones and, from 1994 to 2006, recreational sea
brought to the laboratory in containers with ice, without
urchin harvesting was regulated (50 P. lividus individuals
water, as suggested by Buznikov & Podmarev (1975). Sex
per person per day, during the summer season) in the
determinations were made visually, by puncturing the tes-
tes of freshly dissected specimens and recording, under aLeitz M3C dissection microscope, the presence or absenceof eggs or sperm (Byrne 1990).
Variability in the OSR of P. lividus was estimated over
3 years in July 2004, June 2005 and July 2006, during thesummer sea urchin spawning period (Sa´nchez-Espan˜a
Differences in OSR were analysed using a three-way Anal-
et al. 2004; G. Visconti, unpublished data) at two areas
ysis of Variance (ANOVA) mixed model, with ‘time’ (Ti),
with different density of starfish: the no-take zone A,
three levels (July 2004, June 2005 and July 2006), and
characterized by extremely low presence of Marthasterias
‘Marthasterias density’ (MD), two levels low versus high
glacialis (0.1 ± 0.1 individuals per 250 m2, mean den-
(L versus H), as fixed and orthogonal factors and site (Si),
sity ± SE), and take zone C, characterized by a high pres-
two levels, as a random factor nested in MD. Seven inde-
ence of M. glacialis (3.8 ± 0.5 individuals per 250 m2,
pendent replicates of OSR were used for each combina-
mean density ± SE). Only adult urchins (test diameter
tion of factors for a total of 84 observations. Each
40–45 mm to assure they were matures) were collected by
replicate represents the OSR value estimated within a
SCUBA diving at two sites in the no-take zone A (A1 and
1 m2 quadrat. Data were tested for normality with the
A2, Punta Megna and Acquario, respectively; Fig. 1) and
Bartlett test (Bartlett 1937). Homogeneity of variances
at two sites in the take zone C (C1 and C2, Punta del-
was also checked with Cochran’s C-test (Winer 1971).
l’Arpa and Punta S. Paolo, respectively; Fig. 1). All sites
After ANOVA, means were compared (at a = 0.05) with
were randomly selected, approximately 500 m apart. This
Student–Newman–Keuls (SNK) tests (Underwood 1997).
distance ensured the independence of data: Hereu (2005)
The GMAV 5.0 software (University of Sydney, Australia)
demonstrated that P. lividus has low random mobility on
was used to perform the statistical tests. Furthermore, sig-
rocky bottoms (the total distance observed in a 3-month
nificant deviations of the sex ratio from 0.5 were tested in
study of this species ranged from 71.8 to 673.6 cm). The
each site using a Chi-square test on pooled data across all
study sites were similar in the physical structural com-
replicate quadrats (Sokal & Rohlf 1995).
plexity of their habitat: shallow sublittoral flat basalticgrounds. From the surface to about 3 m depth, the rocky
substrate is generally covered by macroalgae (chiefly artic-ulated Corallinaceae, Dictyotales and Cystoseira spp.) and
sessile invertebrates (e.g. sponges, hydroids, bryozoans).
Thirty M. glacialis, measuring 20–30 cm in diameter (the
From 4 m to about 8 m depth, flat basaltic barren
diameter across the circle defined by the arm tips), were
grounds grazed by sea urchins are present, similar to
randomly collected by SCUBA diving in the study sites
those described in other areas (Frantzis et al. 1988; Sala
from September to October 2007 at a depth of 8 m. After
et al. 1998; Bulleri et al. 1999; Shears & Babcock 2003).
collection, starfishes were placed in large sealed plastic
Overall, apart from different degrees of exposure, there
bags containing oxygenated seawater, for transportation
are no macroscopic differences between locations in terms
to the wet laboratory at the Department of Ecology of the
of rocky substrate, physical heterogeneity, macroalgal
University of Palermo. Starfishes were maintained in 35-l
cover and sea urchin densities: only a small-scale spatial
aquaria with circulating natural seawater, at ambient light
variation (i.e. sites) in term of P. lividus density was
(12 h light and 12 h darkness) and at a temperature of
detected in the present study (F2.108 = 4.42, P = 0.014,
22 °C. Prior to the experiments, all animals were starved
n = 10). At each site, sea urchins were collected along
for a week and kept in separate glass vessels to ensure
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
that all individuals had experienced similar conditions
individualsÆm)2, mean ± SE). Average female density was
and to prevent changes in natural feeding responses due
maximum in July 2006 at site A2 (3.3 ± 0.6 individu-
to the stress of collection and transfer (Rochette et al.
alsÆm)2, mean ± SE) and minimum in June 2005 at site
C1 (0.1 ± 0.1 individualsÆm)2, mean ± SE, Fig. 2).
The laboratory experiment consisted in simultaneously
A male-biased OSR was detected at the sites with high
offering a male and a female P. lividus (test diameter
Martasterias glacialis density but not at those with low
40–45 mm) to a M. glacialis. Sea urchins were sexed
starfish density. Chi-squared results showed a male-
before starting observations by injecting with a syringe
skewed OSR at site C2 in July 2004, at site C1 in June
about 0.1–0.2 ml of KCl 0.5 m into the sea urchin body
2005 and in both C sites in July 2006. In contrast, a
cavity and recording the presence or absence of eggs or
balanced equilibrium between sexes was detected at all
sperm: females produce conspicuous orange eggs, males
times at sites with low M. glacialis density (Table 1).
produce a milky white colour sperm. The injected female
The proportion of male sea urchins ranged from 44.5%
and male P. lividus, with egg and sperm appearing on the
(A2 – June 2005) to 94% (C1 – June 2005) with a
top of the aboral side, were simultaneously placed at the
mean ± SE of 65.5 ± 5.7%, whereas that of females ran-
opposite corners of the experimental aquaria and the
ged from 55.5% (A2 – June 2005) to 6.0% (C1 – June
M. glacialis attack was recorded. We considered an attack
2005) with a mean ± SE of 35.9 ± 6.0%. Mean OSR
successful when we subsequently observed the following
reached its maximum at site C1 in July 2006 (0.85 ± 0.05,
starfish activity: ‘moving toward prey’, ‘capturing prey’
mean ± SE) and minimum in the same year and month
(wrapping its arms around the prey) and ‘digesting prey’(extra-oral digestion with the stomach evaginated between
Table 1. Males and females of Paracentrotus lividus (test diameter
the spines of the sea urchin). Because this last activity
40–45 mm) recorded at the four studied sites in July 2004, June 2005
lasted up to 20–24 h, feeding observations stopped when
and July 2006 and Yates corrected Chi-squared and P-values.
this started. To ensure the independence of data, no M. glacialis specimens were used more than once (Under-
wood 1997). Data from the experiment were analysed
usin the Chi-square goodness-of-fit test, considering as
null hypothesis the sea stars’ lack preference for either sex
We collected a total of 523 sea urchins: 178 males and
185 females at the protected sites (A) and 134 males and
26 females at the harvested ones (C). Average Paracentro-
tus lividus density was maximum in July 2006 at site A
(1 ± 0.1 individualsÆm)2, mean ± SE) in June 2005 at site
1 (Fig. 2). Average male density was maximum in July
2006 at site A1 (4 ± 0.5 individualsÆm)2, mean ± SE)and minimum in July 2004 at site C
*, p < 0.05, ***, p < 0.001; n.s., not significant.
Pooled and separate males and females of Paracentrotus lividus
Fig. 2. Pooled and separate male and female
Paracentrotus lividus density (mean ± SE) atthe four studied sites in July 2004, June 2005
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
Fig. 3. OSR (mean ± SE) of Paracentrotus
lividus (test diameter 40–50 mm) at the four
studied sites in July 2004, June 2005 and July
Table 2. Outcome of the ANOVA on the effects of ‘time’ (July 2004
indicated that the female sex was more vulnerable and
versus June 2005 versus July 2006), ‘Marthasterias density’ (high ver-
that probably eggs were more attractive than sperm to
sus low) and ‘site’ (two level) factors on OSR of Paracentrotus lividus
M. glacialis, suggesting a different mortality between sexes
Many species encounter spatial variation in OSR (Rohr
et al. 2005). A male-biased OSR can be primarily caused
1 Different mating behaviour between sexes. For exam-
ple, the OSR of the blue swimmer crab Portunus pelagicus
(L.) is male-biased during the breeding season because
females seem to require a sandy substrate to extrude their
Data in bold are discussed in the text.
eggs successfully and attach them to the pleopods (Xiao
& Kumar 2004). Similarly, a male-biased OSR wasobserved during the reproductive period in the echinoid
Lytechinus variegatus (Lamarck) at shores of Virginia Key
(0.36 ± 0.06, mean ± SE) (Fig. 3). ANOVA
results revealed significant differences in ‘Marthasterias
in Biscayne Bay; sex-specific aggregation and migration
density’ only (P = 0.004, Table 1) with greater OSR
have been invoked as possible explanations (McCarthy &
values at the sites with high M. glacialis density. No statis-
tical differences were detected for the factor ‘Time’
investment between the two sexes may also bias the OSR
(Bateman 1948). For example in the crab Uca crenulata(Lockington), females typically invest more in each off-spring than males and thus may take longer to recover
from producing offspring and will be available for mating
In the laboratory experiments, 93.3% of available starfish-
less often than mature males (de Rivera 2003).
es made successful attacks. In particular, 20 of 28
Different levels of natural predation on females than
M. glacialis attacked P. lividus females significantly more
on males (de Rivera 2003). For example, a male-biased
often (v2 = 5.1; df = 1; P < 0.02; Fig. 4). Results clearly
OSR is very common in the crab U. crenulata as avianpredators typically prefer the easier-to-handle females(Bildstein et al. 1989).
Potential predators of Paracentrotus lividus in Mediter-
ranean sublittoral rocky habitats include large crustaceans,
asteroids, gastropods and fishes (Sala et al. 1998).
However, only the seabreams Diplodus sargus (L.) and
Diplodus vulgaris (E. Geoffroy St.-Hilaire 1817) have been
reported to effectively control sea urchin populations
(Sala & Zabala 1996; Guidetti 2004).
However, the importance of starfish in regulating Med-
iterranean sea urchin populations has been and is still a
Fig. 4. Feeding preferences of Marthasterias glacialis (n = 28).
matter of debate. According to Fanelli et al. (1999),
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
Marthasteria glacialis may play a keystone role in regulat-
protected sites due to their higher energy content. This
ing abnormal populations of P. lividus, whereas field
could encourage M. glacialis to prefer females to males:
observations made by Guidetti (2004) revealed that this
the effort required to handle and digest this particularly
starfish preyed only occasionally upon P. lividus and
well-defended prey (Guidetti & Mori 2005) would be
Arbacia lixula, suggesting that it is probably unable to
optimized by choosing the individual with higher energy
content. In doing so, M. glacialis may maximize its energy
The present study has been conducted at a place where
gain and minimize the total energetic cost of foraging.
densities of the above-mentioned seabreams are histori-
However, we recognize that detailed field experiments,
cally low (La Mesa & Vacchi 1999) and consequently fish
using a manipulative approach, are needed to confirm the
predation should not exert an effective control on the
role of M. glacialis in controlling the OSR of P. lividus.
abundance of sea urchins (Guidetti & Sala 2007; Guidetti
Other factors, such as differences in growth or mortality
et al. 2008). On the other hand, a high number of
between sexes (McPherson 1965), and different reproduc-
M. glacialis has been observed predating upon P. lividus,
tive behaviour between sexes (i.e. spawning behaviour
in particular in the take area C of the MPA (Bonaviri
with different timing of gamete release, different seasonal
2007). The natural conditions of the MPA of Ustica Island
sex migration and seasonal aggregation), which varies
offered an opportunity to improve our knowledge of the
above all at different density and habitat conditions (Ber-
role of starfish predation in regulating population and
nard 1977; Levitan 2004), may also explain the observed
OSR of sea urchins, especially when fish predation is low.
The results of the present study provided evidence that
This study confirms that regulation of sea urchin den-
the male-skewed OSR observed in July 2002 was real and
sity is a complex and unpredictable process. In fact, in
consistent over time, and it is reasonable to assume that
temperate coastal areas (for reviews, see Sala et al. 1998
this could be caused by a sex-selective predation process.
and Pinnegar et al. 2000) several factors are known to
Laboratory experiment outcomes clearly showed that
control sea urchin population density; among these it is
P. lividus females were more vulnerable than males to
possible to distinguish both extrinsic factors such as epi-
predation by M. glacialis, suggesting a different mortality
demic diseases, food availability, complexity of the sub-
between sexes during the spawning event. This result
strate and availability of refuges, and intrinsic factors
could explain the observed P. lividus OSR pattern at the
such as predation, recruitment, early mortality, migration
MPA of Ustica Island. A male-skewed OSR was found at
rates, etc. (Tegner & Dayton 1977; Jangoux 1987; McCl-
the sites with a high M. glacialis density, whereas a bal-
anahan & Shafir 1990; Sala & Zabala 1996; Lo`pez et al.
anced OSR was observed at sites with a low M. glacialis
1998; Shears & Babcock 2003; Tuya et al. 2005; Pais
et al. 2007). As regards the predation factor, based on
Several studies have tried to identify which elements
our field and laboratory observations, we carefully
make prey more vulnerable to starfish predators and
hypothesize that starfish predation on P. lividus is a
waterborne odours from prey have been proposed as the
process that may be important in regulating population
most important factor in determining predation (Jangoux
& Lawrence 1982; Rochette et al. 1994; Himmelman et al.
To summarise, this preliminary study confirmed the
2005). We showed that the response of M. glacialis to
importance of the sex-selective predation process in influ-
female P. lividus specimens was strong and this is proba-
encing the OSR of a species and in particular provided
bly due to different chemical stimuli related to gamete
important avenues for future work on the role of
M. glacialis in regulating Mediterranean sea urchin popu-
Furthermore, laboratory observations provided evi-
dence that at Ustica MPA the overall gonadosomaticindex (GSI) of P. lividus (test diameter 40–45 mm) dur-
ing the summer spawning period (June 2007) differedwith sex and sea urchin density. GSI was significantly
The authors are indebted to Dr T. Vega Fernandez,
higher in females at the non-protected sites (C area),
Prof. D. Levitan, Dr G. Fanelli, Dr P. Galeotti and two
under low density conditions and low intra-specific com-
anonymous reviewers for their constructive and challeng-
petition for habitat and resources, whereas no differences
ing comments, which substantially improved this manu-
were found at the protected sites (A area, high sea urchin
script; Helen Main for revising the English; the Harbour
density). GSI was also higher in the non-protected sites
Office of Palermo and the MPA management body of Us-
for females, whereas no differences were detected for
tica Island for assistance. This study was funded by the
males (P. Gianguzza, unpublished data). Thus, higher
M.A.T.T.M. (Ministero dell’Ambiente Difesa del Territo-
predation on P. lividus females could be expected at non-
rio e del Mare) research project ‘Monitoraggio delle
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
popolazioni di Paracentrotus lividus e Arbacia lixula ai fini
Frantzis A., Berthon J.F., Maggiore F. (1988) Relation trophi-
della tutela della diversita` biologica dell’ Area Marina
que entre les oursins Arbacia lixula et Paracentrotus lividus
Protetta Isola di Ustica. This paper is dedicated to Mario,
(Echinoidea Regularia) et le phytobenthos infralittoral
the little baby of P.G. who recently came into the world.
superficiel de la baie de Port-Cros (Var, France). ScientificReport of Port-Cros National Park, 14, 81–140.
Gianguzza P., Chiantore M., Bonaviri C., Cattaneo-Vietti R.,
Vielmini I., Riggio S. (2006) The effects of recreational
Bartlett M.S. (1937) Properties of sufficiency and statistical
Paracentrotus lividus fishing on distribution patterns of sea
tests. Proceedings of the Royal Society of London. Series A,
urchins at Ustica Island MPA (Western Mediterranean,
Italy). Fisheries Research, 81, 37–44.
Bateman A.J. (1948) Intra-sexual selection in Drosophila.
Guidetti P. (2004) Consumer of sea urchins, Paracentrotus livi-
dus and Arbacia lixula, in shallow Mediterranean rocky
Bernard F.R. (1977) Fishery and reproduction cycle of the red
reefs. Helgoland Marine Research, 58, 110–116.
sea urchin, Strongylocentrotus franciscanus, in British Colum-
Guidetti P., Mori M. (2005) Morpho-functional defences of
bia. Journal of the Fisheries Research Board of Canada, 34,
Mediterranean sea urchins, Paracentrotus lividus and
Arbacia lixula, against fish predators. Marine Biology, 147,
Bildstein K.L., McDowell S.G., Brisbin I.L. (1989) Conse-
quences of sexual dimorphism in sand fiddler crabs, Uca
Guidetti P., Sala E. (2007) Community-wide effects of marine
pugilator, differential vulnerability to avian predator. Animal
reserves in the Mediterranean Sea. Marine Ecology Progress
Bonaviri C. (2007) Distribuzione delle popolazioni di Paracen-
Guidetti P., Milazzo M., Bussotti S., Molinari A., Murenu
trotus lividus ed Arbacia lixula nell’infralitorale dell’Area Mar-
M., Pais A., Spano N., Balzano R., Agardy T., Boero F.,
ina Protetta ‘‘Isola di Ustica’’ e loro influenza sulla comunita`
Carrada G., Cattaneo-Vietti R., Cau A., Chemello R.,
bentonica. Ph.D Thesis, Universita` degli Studi di Palermo.
Greco S., Manganaro A., Notarbartolo di Sciara G., Russo
Boudouresque C.F., Verlaque M. (2001) Ecology of Paracentro-
F.G., Tunesi L. (2008) Italian marine reserve effectiveness:
tus lividus. In: Lawrence J.M. (Ed.), Edible Sea Urchins.
does enforcement matter? Biological Conservation, 141,
Bulleri F., Benedetti-Cecchi L., Cinelli F. (1999) Grazing by the
Hereu B. (2005) Movement patterns of the sea urchin Paracen-
sea urchin Arbacia lixula L. and Paracentrotus lividus Lmk in
trotus lividus in a marine reserve and an unprotected area in
the Northwest Mediterranean. Journal of Experimental Mar-
the NW Mediterranean. Marine Ecology an evolutionary
ine Biology and Ecology, 241, 81–95.
Buznikov G.A., Podmarev V.K. (1975) The sea urchins Strong-
Himmelman J.H., Dutila T.C., Gaymer C.F. (2005) Foraging
ylocentrotus dro¨bachiensis, S. nudus, and S. intermedius. In:
behaviour and activity budgets of sea stars on a subtidal
Detlaf T.A. (ed), Objects of Developmental Biology (Russ.),
sediment bottom community. Journal of Experimental Mar-
ine Biology and Ecology, 322, 153–165.
Byrne M. (1990) Annual reproductive cycles of the commercial
Jangoux M. (1987) Diseases of Echinodermata. I. Agents
sea urchin Paracentrotus lividus from an exposed intertidal
microorganisms and protistans. Diseases of Aquatic Organ-
and sheltered subtidal habitat on the west coast of Ireland.
Jangoux M., Lawrence J.M. (1982). Echinoderm Nutrition. A.
Clutton-Brock T.H., Parker G.A. (1992) Potential reproductive
rates and the operation of sexual selection. The Quarterly
La Mesa G., Vacchi M. (1999) An analysis of the coastal fish
assemblages of the Ustica Island Marine Reserve (Mediterra-
Clutton-Brock T.H., Vincent A.C.J. (1991) Sexual selection
nean Sea). PSZN: Marine Ecology, 20, 147–165.
and the potential reproductive rates of males and females.
Lawrence J.M. (1987) A Functional Biology of Echinoderms.
Emlen S.T., Oring L.W. (1977) Ecology, sexual selection, and
Lessios H.A. (1979) Use of Panamanian sea urchins to test the
the evolution of mating systems. Science, 197, 215–223.
molecular clock. Nature, 280, 599–601.
Fanelli G., Piraino S., Esposito L., Boero F. (1999) Opposite
Levitan D.R. (2004) Density-dependent sexual selection in
role of sea urchins and starfishes in marine benthic commu-
external fertilizers: variances in male and female fertilization
nities. In: Candia Carnevali N.D., Bonassoro F. (Eds), Echi-
success along the continuum from sperm limitation to sex-
noderm Research. Balkema Publishing, Rotterdam: 453–457.
ual conflict in the sea urchin Strongylocentrotus franciscanus.
Fenberg F.B., Kaustuv R. (2008) Ecological and evolutionary
The American Naturalist, 164, 298–309.
consequences of size-selective harvesting: how much do we
Lipani C., Vitturi R., Sconzo G., Barbata G. (1996) Karyotype
know? Molecular Ecology, 17, 209–220.
analysis of the sea urchin Paracentrotus lividus
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
(Echinodermata): evidence for a heteromorphic chromo-
NW Mediterranean. Marine Ecology Progress Series, 140,
some sex mechanism. Marine Biology, 127, 67–72.
Lo`pez S., Turon X., Montero E., Palacin C., Duarte C.M.,
Sala E., Boudouresque C.F., Harmelin-Vivien M. (1998) Fishing,
Tarjuelo I. (1998) Larval abundance, recruitment and early
trophic cascades, and the structure of algal assemblages:
mortality in Paracentrotus lividus (Echinoidea). Interannual
evaluation of an old but untested paradigm. Oikos, 82,
variability and plankton–benthos coupling. Marine Ecology
Sa´nchez-Espan˜a A.I., Martı´nez-Pita I., Garcı´a F.J. (2004) Gona-
McCarthy D.A., Young M.C. (2002) Gametogenesis and repro-
dal growth and reproduction in the commercial sea urchin
ductive behaviour in the echinoid Lytechinus variegatus.
Paracentrotus lividus (Lamarck, 1816) (Echinodermata: Echi-
Marine Ecology Progress Series, 233, 157–168.
noidea) from Southern Spain. Hydrobiologia, 519, 61–72.
McClanahan T.R., Shafir S.H. (1990) Causes and consequences
Savy S. (1987a) Activity pattern of the sea-star, Marthasterias
of sea urchin abundance and diversity in Kenyan coral reef
glacialis, in Port-Cros Bay (France, Mediterranean coast).
McPherson B.F. (1965) Contributions to the biology of the sea
Savy S. (1987b) Les predateurs de Paracentrotus lividus (Echino-
urchin Tripneustes ventricosus. Bulletin of Marine Sciences,
dermata). France Gis Posidonie Publications, Marseille:
McPherson B.F. (1968) Contributions to the biology of the sea
Shears N.T., Babcock R.C. (2003) Continuing trophic cascade
urchin Eucidaris tribuloides (Lamarck). Bulletin of Marine
effects after 25 years of no-take marine reserve protection.
Marine Ecology Progress Series, 246, 1–16.
Pais A., Chessa L.A., Serra S., Ruiu A., Meloni G., Donno Y.
Sokal R.R., Rohlf F.J. (1995) Biometry: The Principles and Prac-
(2007) The impact of commercial and recreational harvest-
tice of Statistics in Biology Research, 3rd edn. Freeman, New
ing for Paracentrotus lividus on shallow rocky reef sea urchin
communities in North-Western Sardinia, Italy. Estuarine,
Tegner M.J., Dayton P.K. (1977) Sea urchin recruitment pat-
Coastal and Shelf Science, 73, 589–597.
terns and implications of commercial fishing. Science, 196,
Pearse J.S., Cameron R.A. (1991) Echinodermata: Echinoidea.
In: Giese A.C., Pearse J.S., Pearse V.B. (Eds), Reproduction of
Tuya F., Jerez P.S., Haroun R.J. (2005) Influence of fishing
Marine Invertebrates. Vol. VI. Echinoderms and Lophoph-
and functional group of algae on sea urchin control of algal
orates. The Boxwood Press, California: 514–662.
communities in the Eastern Atlantic. Marine Ecology Progress
Pinnegar J.K., Polunin N.V.C., Francour P., Badalamenti F.,
Chemello R., Harmelin-Vivien M., Hereu B., Milazzo M.,
Underwood A.J. (1997) Experiments in Ecology: Their Logic
Zabala M., D’Anna G., Pipitone C. (2000) Trophic cascades
Design and Interpretation Using Analysis of Variance. Cam-
in benthic marine ecosystems: lessons for fisheries and pro-
tected-area management. Environmental Conservation, 27,
Valentincic T. (1973) Food finding and stimuli to feeding in
the sea star Marthasterias glacialis. Netherlands Journal of
de Rivera C.E. (2003) Causes of a male-biased operational sex
ratio in the fiddler crab Uca crenulata. Journal of Ethology,
Verlaque M. (1987) Relations entre Paracentrotus lividus
(Lamarck) et le phytobenthos de Mediterranee´ Occidentale.
Rochette R., Hamel J.F., Himmelman J.H. (1994) Foraging
In: Boudoresque C.F. (Ed.), Colloque International sur Para-
strategy of the asteroid Leptasterias polaris: role of prey
centrotus lividus et les Oursins Comestibles. GIS Posidonie
odors, current and feeding status. Marine Ecology Progress
Winer B.J. (1971) Statistical Principles in Experimental Designs,
Rohr J.R., Park D., Sullivan A.M., McKenna M., Propper C.R.,
Madison D.M. (2005) Operational sex ratio in newts: field
Xiao Y., Kumar M. (2004) Sex ratio, and probability of sexual
responses and characterization of a constituent chemical
maturity of females at size, of the blue swimmer crab, Port-
cue. Behavioral Ecology, 16, 286–293.
unus pelagicus Linneaus, off Southern Australia. Fisheries
Sala E., Zabala M. (1996) Fish predation and the structure
of the sea urchin Paracentrotus lividus populations in the
Marine Ecology (2008) 1–8 ª 2008 The Authors. Journal compilation ª 2008 Blackwell Publishing Ltd
Artigos Teoria da Complexidade: Percursos e Desafios para a Pesquisa em Educação Adriana Marques Paderes* Professora das Faculdades de Valinhos - FAVe-mail: [email protected] Regina de Brito Rodrigues Sonia Regina Giusti Especialista em Recursos Humanos na Gestãode Negócios - Faculdades São Judas Tadeu Abstract Reflete-se, neste artigo, sobre os
3 2 n d E D I T I O N Carbapenems: Interaction with sodium valproate Doripenem monohydrate (marketed as Doribax), 4.4 Special warnings and precautions for use a synthetic antibiotic, is a new chemical entity The concomitant use of doripenem and valproic that belongs to the carbapenem class of beta-lac- acid/sodium valproate is not recommended (see section tams. Doripenem is administer
 Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
take zone C (high starfish M. glacialis density and a
particular in the take area of the MPA (authors’ personal
balanced OSR at the no-take zone A (low starfish den-
observation). Unfortunately, due to the extremely cryptic
sity). Given the ecological and economic importance of
behaviour and nocturnal activity of this species (Savy
P. lividus, this operational sex-ratio disparity was consid-
1987b), abundance estimates are problematic and may be
ered worthy of further investigation. The apparent imbal-
biased. However, a high number of M. glacialis have been
ance in the OSR may reflect a sampling procedure bias,
observed predating sea urchins in recent years, and the
but if this pattern is confirmed, this may have important
remains of sea urchins eaten by starfish are frequently
consequences for fluctuations in population size and
recorded, especially at the barren grounds of zone C
structure, especially in a small population (Emlen &
M. glacialis is a slow-moving predator, able to detect
P. lividus is a species with a chromosomal (genetic) sex
its prey using olfaction (Valentincic 1973). It could be
determination mechanism expected to allocate equal
hypothesized that during the P. lividus spawning period
effort to producing male and female offspring because
the starfish is more attracted by (and hence preferentially
random meiosis would lead to a mean sex ratio of 0.5
consumes) females. The aim of this study was to explore
(Lipani et al. 1996). In this case, environmental condi-
possible variability in OSR of P. lividus populations
tions, such as pronounced temperature fluctuations,
inhabiting barren rocky substrates of Ustica Island MPA.
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
take zone C (high starfish M. glacialis density and a
particular in the take area of the MPA (authors’ personal
balanced OSR at the no-take zone A (low starfish den-
observation). Unfortunately, due to the extremely cryptic
sity). Given the ecological and economic importance of
behaviour and nocturnal activity of this species (Savy
P. lividus, this operational sex-ratio disparity was consid-
1987b), abundance estimates are problematic and may be
ered worthy of further investigation. The apparent imbal-
biased. However, a high number of M. glacialis have been
ance in the OSR may reflect a sampling procedure bias,
observed predating sea urchins in recent years, and the
but if this pattern is confirmed, this may have important
remains of sea urchins eaten by starfish are frequently
consequences for fluctuations in population size and
recorded, especially at the barren grounds of zone C
structure, especially in a small population (Emlen &
M. glacialis is a slow-moving predator, able to detect
P. lividus is a species with a chromosomal (genetic) sex
its prey using olfaction (Valentincic 1973). It could be
determination mechanism expected to allocate equal
hypothesized that during the P. lividus spawning period
effort to producing male and female offspring because
the starfish is more attracted by (and hence preferentially
random meiosis would lead to a mean sex ratio of 0.5
consumes) females. The aim of this study was to explore
(Lipani et al. 1996). In this case, environmental condi-
possible variability in OSR of P. lividus populations
tions, such as pronounced temperature fluctuations,
inhabiting barren rocky substrates of Ustica Island MPA.

 Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
Fig. 3. OSR (mean ± SE) of Paracentrotus
lividus (test diameter 40–50 mm) at the four
studied sites in July 2004, June 2005 and July
Table 2. Outcome of the ANOVA on the effects of ‘time’ (July 2004
indicated that the female sex was more vulnerable and
versus June 2005 versus July 2006), ‘Marthasterias density’ (high ver-
that probably eggs were more attractive than sperm to
sus low) and ‘site’ (two level) factors on OSR of Paracentrotus lividus
M. glacialis, suggesting a different mortality between sexes
Many species encounter spatial variation in OSR (Rohr
et al. 2005). A male-biased OSR can be primarily caused
1 Different mating behaviour between sexes. For exam-
ple, the OSR of the blue swimmer crab Portunus pelagicus
(L.) is male-biased during the breeding season because
females seem to require a sandy substrate to extrude their
Data in bold are discussed in the text.
Gianguzza, Badalamenti, Gianguzza, Bonaviri & Riggio
Fig. 3. OSR (mean ± SE) of Paracentrotus
lividus (test diameter 40–50 mm) at the four
studied sites in July 2004, June 2005 and July
Table 2. Outcome of the ANOVA on the effects of ‘time’ (July 2004
indicated that the female sex was more vulnerable and
versus June 2005 versus July 2006), ‘Marthasterias density’ (high ver-
that probably eggs were more attractive than sperm to
sus low) and ‘site’ (two level) factors on OSR of Paracentrotus lividus
M. glacialis, suggesting a different mortality between sexes
Many species encounter spatial variation in OSR (Rohr
et al. 2005). A male-biased OSR can be primarily caused
1 Different mating behaviour between sexes. For exam-
ple, the OSR of the blue swimmer crab Portunus pelagicus
(L.) is male-biased during the breeding season because
females seem to require a sandy substrate to extrude their
Data in bold are discussed in the text.