Examining neurochemical determinants of inspection time
Con Stougha,*, J.C. Thompsonb, T.C. Batesc, P.J. Nathand
aNeuropsychology Laboratory, BSEE, Swinburne University, P.O. Box 218, Hawthorn, VIC 3122, Australia
bBrain Sciences Institute, Swinburne University, Hawthorn, VIC 3122, Australia
cDepartment of Psychology, Macquarie University, North Ryde, NSW, Australia
dNeuropharmacology Laboratory, Swinburne University, Hawthorn, VIC 3122, Australia
Received 13 March 2001; accepted 13 March 2001
Inspection time (IT), an information-processing correlate of psychometric intelligence, has been
extensively studied. Previous research has shown that IT is a reliable correlate of psychometricintelligence across different developmental periods, mirroring developmental trends of fluidintelligence. Despite this extensive previous literature, very little is known about the biological basisof IT. In the present review, we discuss recent results from our laboratories examining theneurochemical determinants of IT. In this review, we outline the significance of several studies inwhich performance on the IT task is measured before and after modulating key human central nervoussystem (CNS) neurotransmitters and receptor systems (e.g., cholinergic, serotonergic, noradrenergic,and dopaminergic systems). The results of these studies indicate a primarily cholinergic basis for IT,although other aspects of psychometric intelligence may have serotonergic and dopaminergicdeterminants in addition to a cholinergic basis. The results are consistent with data reportingcholinergic depletion and impaired IT performance in dementia of the Alzheimer’s type (DAT). Speculatively, we propose that compounds that enhance the release of the neurotransmitteracetylcholine (Ach) will improve IT and the variance that IT shares with IQ test performance. D 2001 Elsevier Science Inc. All rights reserved.
Keywords: Inspection time; Intelligence; Acetylcholine; Pharmacology
* Corresponding author. Tel.: +61-3-9214-8167; fax: +61-3-9214-5230. E-mail address: [email protected] (C. Stough).
0160-2896/01/$ – see front matter D 2001 Elsevier Science Inc. All rights reserved. PII: S 0 1 6 0 - 2 8 9 6 ( 0 1 ) 0 0 0 7 7 - 0
C. Stough et al. / Intelligence 29 (2001) 511–522
Visual inspection time (IT) is defined as the minimum time an individual requires to reach
near perfect response accuracy in a simple two-choice visual discrimination. Manipulating thetime between stimulus onset and the onset of a backward mask controls stimulus duration. The backward mask acts to prevent further iconic sampling of the stimulus (Vickers,Nettelbeck, & Willson, 1972). IT is not dependent on response speed, and as such is notconfounded by changes to motor speed. IT is widely regarded as a measure of the early stagesof information processing (Deary & Stough, 1996). IT has been shown to correlate reliablywith performance on standard tests of psychometric intelligence (Deary & Stough, 1996,1997; Stough et al., 1996) and has been proposed as an experimental model for understandingthe biological basis of a component of human intelligence. Along these lines, there have beenmuch research describing developmental changes in IT across the lifespan that appear toapproximate changes in fluid intelligence (Anderson, 1989; Nettelbeck & Young, 1989,1990). Several studies have examined both empirical and theoretical relationships between ITand the cognitively more complex IQ, with reviews and meta-analyses concluding thataround 25% of the psychometric IQ variance can be reliably accounted for by IT (Deary &Stough, 1996; Kranzler & Jensen, 1989; Nettelbeck, 1987). Such findings, in addition to thetheory that differences in IQ may be a result of individual differences in mental speed, haveled to suggestions that IT may be a biological or semibiological measure of intelligence(Deary & Stough, 1996).
Despite this previous research, there has been little research examining the biological
correlates or causes of IT. Deary and Caryl (1997) have previously discussed studiesexamining the electrophysiological correlates of IT and there have been some recentresearch (published in the current volume of Intelligence) describing fMRI correlates ofIT performance (Deary, Simonotto, Marshall, Goddard, & Wardlaw, 2001). Data from brainimaging research allow hypotheses to be evaluated ascribing the influence of differentneural regions on IT performance. However, brain imaging data, particularly at thisrelatively early stage of research, do not allow us to infer the events at a neuronal levelunderpinning IT performance. A complete biological model of IT, or for that matter themore cognitively complex IQ, requires an understanding at both the brain structural and atthe neurochemical level. It is also likely that an understanding of neurochemical processes(at the level of the neuron) may facilitate an understanding of the genetic mechanisms ofboth IT and IQ as receptors and neurotransmitters are intimately involved in gene regulation(Stahl, 2000). Pharmacological agents that act on receptors that underlie the secondarymessenger system send signals that ultimately regulate the expression of our genes. Thepresent article reviews recent work primarily from our own laboratories, in which we assessthe relative contribution of several key human neurotransmitters on IT performance in
C. Stough et al. / Intelligence 29 (2001) 511–522
healthy participants. We outline our theory and empirical support for a predominantlycholinergic basis for IT.
2. Acetylcholine, cholinergic neurotransmission, and IT
2.1. Early work on the pharmacology of IT: nicotine
Work in this area first developed from our observation that nicotine enhanced both
intelligence test performance (Stough, Mangan, Bates, & Pellett, 1994) and information-processing correlates of IQ such as DT from the Jensen RT paradigm (Bates, Pellett, Stough,& Mangan, 1994), the string length of the auditory evoked potential (AEP; Stough, Bates,Mangan, & Pellett, 1995), and crucially for present considerations, IT (Stough et al., 1995). Attempting to understand this effect led us to propose that IT and indeed the variance that ITshares with IQ was predominantly mediated by cholinergic neurotransmission (Stough et al.,1996; Thompson, Stough, Nathan, Ames, & Ritchie, 2000). Fig. 1 illustratively outlines thecholinergic system in which presynaptic nicotine receptors, when activated by a suitableagonist (e.g., nicotine), release the neurotransmitter acetylcholine (Ach) across the synapse
Fig. 1. Diagrammatic representation of the cholinergic neurotransmitter system: presynaptic neuron, synapse,postsynaptic neuron, cholinergic receptors (both muscarinic and nicotinic), acetylcholinesterase inhibitor, and theneurotransmitter Ach.
C. Stough et al. / Intelligence 29 (2001) 511–522
(i.e., presynaptic release). This increase in Ach increases the probability of the neuro-transmitter, finding a suitable postsynaptic receptor (in the case of the cholinergic system, thisis predominantly muscarinic receptors but there is also limited postsynaptic nicotinicreceptors). Fig. 1 also illustrates the key components of the cholinergic system includingneurotransmitter (Ach), pre- and postsynaptic receptors (nicotinic and muscarinic), andenzyme that metabolises Ach. In Fig. 1, Ach is released from presynaptic vesicles (middleof Fig. 1). It should probably also be pointed out here that the neurochemical approach thatwe have employed is to systematically manipulate the various elements of this system and toobserve the effect on IT performance. In all studies, we have employed placebo-controlleddouble-blind methodologies. Fig. 2 illustratively describes the mechanism by which theneurotransmitter Ach and acetylcholinesterase inhibitor act within this system. This is a keyelement of this system and will be discussed in more detail later.
Consistent with our early work assessing the effects of nicotine on intelligence test
performance and on biological correlates of intelligence test performance, there is consid-erable evidence suggesting that acute nicotine enhances speed of information processing andvigilance, as well as other cognitive processes such as attention and memory (Bates, Mangan,Stough, & Corballis, 1995; Stough et al., 1995; for a review, see Warburton, 1992). However,one difficulty in ascribing this nicotine-related cognitive enhancement to the effects ofnicotine receptor activity is that these studies were conducted on smoking subjects, usuallyafter a 2-h or longer period of nicotine abstinence. This is a consistent methodologicalproblem in many studies examining the effects of nicotine on human performance — fewstudies have examined nicotine administration in nonsmokers whose performance cannot be
Fig. 2. Diagrammatic representation of the action of acetylcholinesterase inhibitors on the neurotransmitter Ach. Acetylcholinesterase inhibitors prevent the breakdown of the neurotransmitter increasing the concentration andduration of Ach in the synapse increasing the likelihood of postsynaptic activation. Postsynaptic activation ofcholinergic neurons is hypothesised to mediate IT.
C. Stough et al. / Intelligence 29 (2001) 511–522
ascribed to the amelioration of nicotine withdrawal. In order to assess this potentialmethodological problem, two recent studies from our laboratories have examined the effectsof non-cigarette administration of nicotine in nonsmokers on IT performance. In the firststudy by Stough, Wilby, and Thompson (submitted for publication), nonsmokers recordedsignificantly shorter ITs (i.e., faster information processing) from nicotine patch adminis-tration than from the no-nicotine condition. This effect was greatest at 6 h post-nicotine patchadministration, commensurate with the peak nicotine plasma levels for cutaneous admin-istration of nicotine. The size of the effect approximated the effect shown by Stough et al. (1995) in which nicotine was administered via cigarettes. Interestingly, there was also asignificant shortening of IT at 3 h although this was less than at 6 h, again approximating therelative nicotine plasma levels at this time (Pomerleau & Pomerleau, 1992). In a subsequentstudy, Pyman, Nathan, Thompson, and Stough (submitted for publication) also observed asignificant shortening in IT following nicotine administration from gum in nonsmokers. Boththese studies suggest that IT is modulated by nicotinic receptor activity independent of anyamelioration of nicotine withdrawal. That is, nicotine is a cognitive-enhancing drug, with itsadministration activating presynaptic nicotinic receptors. Nicotine receptors are ligand-boundreceptors in which Ca + ions are released into the cell. This causes a depolarisation of thepresynaptic neuron leading to the release of the neurotransmitter Ach (Stahl, 2000).
Consolidating evidence for a role of nicotinic receptors also emerges from studies in which
IT or speed of information processing measures have been assessed in patients with nicotinicAch receptor (nAchRs) loss. McCrimmon, Deary, Huntly, MacLeod, and Frier (1996) reportedthat insulin-induced hypoglycaemia impaired IT, and there is evidence to suggest that insulin-induced impairments to cognition are mediated by nAchRs (Kopf & Baratti, 1999). Patientswith Alzheimer’s disease were reported by Deary, Hunter, Langan, and Goodwin (1991) tohave significantly slower ITs than age- and premorbid IQ-matched Korsakoff’s patients andcontrols. In these patients, IT was correlated — .81 with CAMDEX-Cog scores. Recentresearch by Mackie, Thompson, and Stough (submitted for publication) has also indicated thatIT was the most impaired test of a large battery of neuropsychological tests routinely used todetect cognitive changes in 41 patients with mild to moderate dementia of the Alzheimer’stype (DAT). Loss of nAchR density is suggested to be one of the critical mechanismsunderlying cognitive impairments in Alzheimer’s Disease (Whitehouse & Kalaria, 1995). Such findings suggest that nAchRs are crucial to the neurophysiological basis of IT, andfurthermore suggest that IT may be an index of the integrity of the nAchR system.
2.3. Development of a pharmacological model of IT: other cholinergic components
For such a model to be pharmacologically valid, it is also necessary to show that
antagonism (i.e., blockade) of nicotinic receptors also leads to impairment in IT. In a recentstudy, Thompson et al. (2000) administered the nicotinic receptor antagonist mecamylamineto healthy participants and observed that this acutely impaired IT performance. Interestingly,
C. Stough et al. / Intelligence 29 (2001) 511–522
administration of the cholinomimetic donepezil (acetylcholinesterase inhibitor) partiallyameliorated this impairment in IT suggesting that IT may, at least in part, be mediated bycholinergic processes. Donepezil is a cholinesterase inhibitor, which increases the level ofsynaptic Ach by inhibiting its metabolism by the choline enzyme (Stahl, 2000, see Fig. 2). IfIT is modulated by presynaptic nicotinic receptor activity releasing endogenous Ach forpostsynaptic receptor binding, then donepezil should at least partially ameliorate the blockadeof presynaptic nicotinic receptors by increasing the level of synaptic Ach to bind withpostsynaptic receptors. This was indeed confirmed by Thompson et al. (2000).
Further research from our laboratories has also examined muscarinic blockade and IT
performance. Early studies have suggested that the blockade of muscarinic receptors (hypoth-esised to be primarily active postsynaptically in the human cortex) did not impair IT perform-ance. A study by Parrott and Deary (1992) reported that the muscarinic antagonist scopolaminedid not impair IT. Similarly, Kennedy, Odenheimer, Baltzley, Dunlap, and Wood (1990)reported that scopolamine did not impair measures of perceptual speed. However, our ownresearch suggests that muscarinic blockade does significantly impair IT but unlike nicotinicreceptor blockade, administration of the cholinomimetic donepezil (acetylcholinesteraseinhibitor) did not ameliorate this effect (Waterham, Thompson, Nathan, & Stough, submittedfor publication). A potential explanation for this finding was that the increase in the level ofsynaptic Ach might not have been sufficient to displace scopolamine from muscarinicreceptors. The inconsistent results may in part be due to the different methodologies used inthe three studies, particularly the dose and time of administering the scopolamine. The result ofthis scopolamine study taken together with the results of a recent study by Hutchison, Nathan,Mrazek, and Stough (2001) in which an improvement in IT was observed after globallyincreasing Ach and cholinergic transmission with the cholinomimetic donepezil in healthyparticipants is consistent with a theory that proposes that IT is primarily mediated by synapticlevels of Ach. The action of donepezil is illustrated in Fig. 2. While increasing Ach globallyleads to activation of both nicotinic and muscarinic receptor, we propose the following cascadeof events leading to modulation of IT. Presynaptic release of Ach is controlled by nicotinicreceptors such that enhancing nicotinic receptor function leads to release of Ach. Once released,Ach predominantly activates postsynaptic muscarinic receptors leading to cellular change thattriggers a cascade of events leading to modulation of IT. A post hoc explanation for theWaterham et al. (submitted for publication) observation that donepezil did not ameliorate the ITimpairment from the muscarinic blockade is that Ach would be required to compete withscopolamine for postsynaptic muscarinic receptors. No such difficulty would be observed if themechanism underlying IT was postsynaptic nicotinic receptors as scopolamine is a specificmuscarinic receptor blocker. That is, these studies indicate a role for presynaptic nicotinicreceptors and postsynaptic muscarinic receptors in IT performance.
It is also well known that the low molecular weight amines that are active in the central
nervous system [CNS; such as Ach, serotonin (5-HT), noradrenaline (NA), and dopamine
C. Stough et al. / Intelligence 29 (2001) 511–522
(DA)] have a modulatory effect on each other. That is, elevating the level of one neuro-transmitter may cause an increase or a decrease in the level of another neurotransmitter. Nicotine may, in addition to increasing levels of Ach synaptically, also lead to an increase inother key neurotransmitters, particularly DA. In order to rule out the effects of otherneurotransmitters on IT, we have recently carried out two studies specifically studying therole of monoamines — 5-HT, DA, and NA — on IT performance. These studies offer onlypart of the potential story as we have not yet systematically studied all of the components ofthese neurotransmitter systems (i.e., agonist and antagonists on all receptor classes withineach system). Importantly, DA, 5-HT, and NA have been previously implicated in human andanimal cognitive performance. The neurotransmitters 5-HT and NA are important modulatorsof human cognitive processes such information processing, attention, and working memory(Coull, 1998; Hasbroucq, Rihet, Blin, & Possamai, 1997; Hindmarch, 1981). Impairments inattention and working memory have been reported after attenuation of noradrenergic functionwith the a2 agonist clonidine (see review by Coull, 1998), while an improvement ininformation processing, attention, and memory has been reported following enhancementof noradrenergic function (Clark, Geffen, & Geffen, 1989; Smith, Wilson, Glue, & Nutt,1992). Similarly, enhancement of serotonergic function has been shown to improve sustainedattention, and efficiency of information processing (Hindmarch & Bhatti, 1988; Hindmarch &Harrison, 1988; Lader et al., 1986; Nathan, Stough, & Siteram, 2000). Previc (1999) suggeststhat DA is a key neurotransmitter in the regulation of human intelligence. DA is found in highconcentrations in the dorsolateral, prefrontal, and anterior cingulate cortices (Gaspar, Berger,Febvret, Vigny, & Henry, 1989), areas that are believed to play an important role in thefunction of working memory, mental effort, and the cognitive ‘‘executive’’ (Haut, Kuwabara,Leach, & Callahan, 2000; Kessels, Postma, Wijnalda, & de-Haan, 2000). Previous researchhas revealed that DA depletion in these areas of the brain could be a major contributing factorin most prominent disorders with psychological or neurological symptoms (Comings et al.,1996), including Alzheimer’s disease and other cognitive disorders of aging (Arnsten &Goldman-Rakic, 1985; Ollat, 1992; Volkow, Gur, Wang, & Fowler, 1998). It has been shownthat DA neurons (mainly the DA1 receptor) in the principal sulcus region of the prefrontalcortex play an important role in the working memory of monkeys (Sawaguchi & Goldman-Rakic, 1997; Sawaguchi, Matsumura, & Kubota, 1990). Luciana, Collins, and Depue (1998)showed that the DA agonist bromocriptine improves working memory function in humansparticularly for those with average or below average IQs. Similar results were reported byKimberg, Esposito, and Farah (1997), who showed that the effects of bromocriptine wasdependent on working memory capacity.
3.1. Serotonin, noradrenaline, and dopamine
In the first study designed to test the potential contribution of these other neurotransmitters
on IT performance, Stough, Sitaram, and Nathan (submitted) administered venlafaxine (aselective 5-HT and NA reuptake inhibitor) and citalopram (a selective 5-HT reuptakeinhibitor) to healthy participants. Venlafaxine increases the synaptic levels of both 5-HTand NA, whereas citalopram increases the synaptic level of 5-HT. The results of this double-
C. Stough et al. / Intelligence 29 (2001) 511–522
blind placebo-controlled study indicated that there was not a significant improvement in ITrelative to placebo attributable to either the venlafaxine or citalopram treatment. Furtherresearch employing systematic manipulations of these systems however are required beforewe will be able to exclude any contribution for 5-HT or NA on IT.
Interestingly, a similar study by the same authors (Nathan et al., 2000) showed that there
was a significant change in other cognitive processes due to 5-HT and NA. Relative toplacebo there was a significant change in choice reaction time and critical flicker fusiondue to increasing levels of 5-HT. This result heralds the possibility that intelligence, ifregarded as the sum of many different cognitive processes (e.g., memory, attention, abstractreasoning, spatial ability, perceptual speed, and so on), could be determined by differentneurochemical processes. Whereas IT may be primarily due to Ach, attention and problemsolving may be modulated by 5-HT and or other neurotransmitters. This was further testedby a recent study by Stough, Singh, Lloyd, and Nathan (submitted for publication) inwhich the effect of the DA agonist bromocriptine was assessed on IT, digit span, and digitsymbol. These three tests between them assess a relatively wide range of cognitiveprocesses that include perceptual speed, working memory, concentration and attention,freedom from distractibility, motor preparation and execution, verbal reception, and so on. Acute administration of bromocriptine did not significantly alter IT at peak plasma level(1.5 h after administration). However, significant improvement in both digit span and digitsymbol was observed, consistent with previous research in both humans and animalsindicating that DA is an important contributor to intellectual processes, particularlyworking memory.
These studies taken together sketch out an important role for the neurotransmitter Ach and
the cholinergic system in IT and further substantiate the role of other neurotransmitters indetermining other cognitive processes. The relationship between these neurotransmitters andhow they collectively organise the disparate elements of human intelligence is not wellunderstood. Future work examining the neurochemical determinants of different intellectualprocesses is required. Of particular interest may be the neurochemical basis of cognitiveprocesses with differing g loadings.
There is substantial work to be done on developing an adequate neurochemical model of
IT or the more complex IQ. Such systematic manipulation may involve not only selectiveassessment of different receptor types but kinetic work establishing the peak behaviouraleffects of these drugs and other drugs that also effect these systems. Table 1 presents a verypreliminary summary of pharmacological studies that have assessed IT performance. Itshould be noted that at this stage significantly greater research is required to make Table 1 acomprehensive one.
Obviously acute studies examining more than one neurotransmitter system at a time is also
required. Such studies may involve agonising Ach but impairing 5-HT receptors, vice versa,and so on.
C. Stough et al. / Intelligence 29 (2001) 511–522
Table 1Preliminary summary of neurotransmitter – IT interactions
Future research examining both the neurochemistry and brain imaging during the IT task
are clearly required. Indeed, the integration of these two approaches may significantlyenhance our understanding of the biological basis of this key human intelligence correlate. Given our neurochemical data, we hypothesise that areas rich in cortical Ach are likely toplay important roles in terms of areas that are uncovered as active from metabolic brainimaging studies (such as PET and fMRI). It may also be possible to correlate the regionalselective binding of radioactive cholinergic ligands with IT performance, and even imagetask-related release of endogenous neurotransmitters (Kegeles & Mann, 1997). Research inthis area may also provide tools for the early detection of neuropsychiatric disorders that areprimarily due to cholinergic depletion and which has been the focus of a recent commentaryon information processing and neuropsychiatric disorders by Sarter and Bruno (1999). Further, research in this area may contribute to methods enhancing intelligence pharmaco-logically in the general community or in intellectual disability.
Investigation of the molecular basis of IT may benefit from the development of an
animal model of IT. The increasing sophistication and, more importantly, the constructvalidity of recent rodent models of choice reaction time (Robbins, 1998), visuospatialselective attention (Phillips & Brown, 1999) and sustained attention (McGaughy & Sarter,1995) provide reason for optimism that a rodent model of IT may be possible. Such amodel would allow for more rigorous examination of the role of neurotransmitters, andtheir interactions, using receptor subtype-selective agonists and antagonists (e.g., Stitzel,Lu, Jimenez, Tritto, & Collins, 2000), in vivo microdialysis and electrophysiology (e.g.,Gill, Sarter, & Givens, 2000; Tani, Saito, Imoto, & Ohno, 1998), gene knockout (e.g., Lena& Changeux, 1997), and antisense oligonucleotides (e.g., le Novere et al., 1999; Listerud,Brussaard, Devay, Coleman, & Role, 1991).
The present research also raises important questions about our conceptions of intelligence.
Can intelligence be improved pharmacologically? Is subaverage intelligence a consequenceof poor cholinergic activity, loss of receptors, and associated activity? Is cholinergic systemfunctioning a direct consequence of the expression of our genes or as presumed the
C. Stough et al. / Intelligence 29 (2001) 511–522
consequence of an interaction of our genes and events from the environment that signal generegulation. If the latter, then what are the key environmental switches for receptor up- anddown-regulation? Such possibilities await integrative biological research at the level of thegenes, biology, and environment.
Anderson, M. (1989). New ideas in intelligence. Psychologist: Bulletin of the British Psychological Society, 2 (3),
Arnsten, A. F., & Goldman-Rakic, P. S. (1985). Catecholamines and cognitive decline in aged nonhuman pri-
mates. Annals of the New York Academy of Sciences, 444, 218 – 234.
Bates, T., Mangan, G., Stough, C., & Corballis, P. (1995). Smoking, processing speed and attention in a choice
reaction time task. Psychopharmacology, 120, 209 – 212.
Bates, T. C., Pellett, O., Stough, C., & Mangan, G. L. (1994). The effects of smoking on simple and choice
reaction time. Psychopharmacology, 106, 365 – 368.
Clark, C. R., Geffen, G. M., & Geffen, L. B. (1989). Catecholamines and the covert orientation of attention in
humans. Neuropsychologia, 27, 131 – 139.
Comings, D., Wu, S., Chiu, C., Ring, R., Gade, R., Ahn, C., MacMurray, J., Dietz, G., & Muhleman, D. (1996).
Polygenic inheritance of Tourette syndrome, stuttering, attention deficit hyperactivity, conduct and opposi-tional defiant disorder: the additive and subtractive effect of the three dopaminergic genes-DRD2, D beta H,and DAT1. American Journal of Medical Genetics, 67, 264 – 288.
Coull, J. T. (1998). Neural correlates of attention and arousal: insights from electrophysiology, functional neuro-
imaging and psychopharmacology. Progress in Neurobiology, 55, 343 – 361.
Deary, I. J., & Caryl, P. G. (1997). Neuroscience and human intelligence differences. Trends in Neurosciences, 20
Deary, I. J., Hunter, R., Langan, S. J., & Goodwin, G. M. (1991). Inspection time, psychometric intelligence and
clinical estimates of cognitive ability in pre-senile Alzheimer’s disease and Korsakoff’s patients. Brain, 114,2543 – 2554.
Deary, I. J., Simonotto, E., Marshall, A., Goddard, N., & Wardlaw, J. M. (2001). The functional anatomy of
inspection time: a pilot fMRI study. Intelligence, 29, 497 – 510.
Deary, I. J., & Stough, C. (1996). Intelligence and inspection time: achievements, prospects, and problems.
American Psychologist, 51, 599 – 608.
Deary, I. J., & Stough, C. (1997). Looking down on intelligence. American Psychologist, 52, 551 – 552. Gaspar, P., Berger, B., Febvret, A., Vigny, A., & Henry, J. (1989). Catecholamine innervation of the human
cerebral cortex as revealed by comparative immunohistochemistry of tyrosine hydroxylase and dopamine betahydroxylase. Journal of Comparative Neurology, 279, 249 – 271.
Gill, T. M., Sarter, M., & Givens, B. (2000). Sustained visual attention performance-associated prefrontal neuronal
activity: evidence for cholinergic modulation. Journal of Neuroscience, 20 (12), 4745 – 4757.
Hasbroucq, T., Rihet, P., Blin, O., & Possamai, C. (1997). Serotonin and human information processing: fluvox-
amine can improve reaction time performance. Neuroscience Letters, 299, 204 – 208.
Haut, M. W., Kuwabara, H. O., Leach, S., & Callahan, T. (2000). Age-related changes in neural activation during
working memory performance. Aging, Neuropsychology, and Cognition, 7 (2), 119 – 129.
Hindmarch, I. (1981). Measuring the effect of psychoactive drugs on higher brain function. Advances in Human
Hindmarch, I., & Bhatti, J. Z. (1988). Psychopharmacological effects of sertraline in normal, healthy volunteers.
European Journal of Clinical Pharmacology, 35, 221 – 223.
Hindmarch, I., & Harrison, C. (1988). The effects of paroxetine and other antidepressants in combination with
alcohol in psychomotor activity related to car driving. Human Psychopharmacology, 3, 13 – 20.
C. Stough et al. / Intelligence 29 (2001) 511–522
Hutchison, C. W., Nathan, P. J., Mrazek, L., & Stough, C. (2001). Cholinergic modulation of speed of early
information processing: the effect of donepezil on inspection time. Psychopharmacology, 44, 440 – 442.
Kegeles, L. S., & Mann, J. J. (1997). In vivo imaging of neurotransmitter systems using radiolabeled receptor
ligands. Neuropsychopharmacology, 17 (5), 293 – 307.
Kennedy, R. S., Odenheimer, R. C., Baltzley, D. R., Dunlap, W. P., & Wood, C. D. (1990). Differential effects of
scopolamine and amphetamine on microcomputer-based performance tests. Aviation, Space and EnvironmentalMedicine, 61, 615 – 621.
Kessels, R. P. C., Postma, A., Wijnalda, E., & de-Haan, E. H. F. (2000). Frontal lobe involvement in spatial
memory: evidence from PET, fMRI, and lesion studies. Neuropsychology Review, 10 (2), 101 – 113.
Kimberg, D. Y., D’Espisito, M., & Farah, M. J. (1997). Effects of bromocriptine on human subjects depend on
working memory capacity. Neuroreport, 8 (16), 3581 – 3585.
Kopf, S. R., & Baratti, C. M. (1999). Effects of posttraining administration of insulin on retention of a habituation
response in mice: participation of a central cholinergic mechanism. Neurobiology of Learning and Memory, 71,50 – 61.
Kranzler, J. H., & Jensen, A. R. (1989). Inspection time and intelligence: a meta-analysis. Intelligence, 13,
Lader, M., Melhuish, A., Frcka, G., FredricsonOver, K., & Christensen, V. (1986). The effects of citalopram in
single and repeated doses and with alcohol on physiological and psychological measures in healthy subjects. European Journal of Clinical Pharmacology, 31, 183 – 190.
le Novere, N., Zoli, M., Lena, C., Ferrari, R., Picciotto, M. R., Merlo-Pich, E., & Changeux, J. P. (1999).
Involvement of alpha6 nicotinic receptor subunit in nicotine-elicited locomotion, demonstrated by in vivoantisense oligonucleotide infusion. Neuroreport, 10 (12), 2497 – 2501.
Lena, C., & Changeux, J. P. (1997). Role of Ca2+ ions in nicotinic facilitation of GABA release in mouse
thalamus. Journal of Neuroscience, 17 (2), 576 – 585.
Listerud, M., Brussaard, A. B., Devay, P., Colman, D. R., & Role, L. W. (1991). Functional contribution of
neuronal AChR subunits revealed by antisense oligonucleotides. Science, 254, 1518 – 1521.
Luciana, M., Collins, P. F., & Depue, R. A. (1998). Opposing roles for dopamine and serotonin in the modulation
of human spatial working memory functions. Cerebral Cortex, 8, 218 – 226.
Mackie, D., Thompson, & Stough, C. (submitted for publication). Deficits in the speed of early information
processing speed in dementia of the Alzheimer’s type.
McCrimmon, R. J., Deary, I. J., Huntly, B. J. P., MacLeod, K. J., & Frier, B. M. (1996). Visual information
processing during controlled hypoglycaemia in humans. Brain, 119, 1277 – 1287.
McGaughy, J., & Sarter, M. (1995). Behavioral vigilance in rats: task validation and effects of age, amphetamine,
and benzodiazepine receptor ligands. Psychopharmacology, 117 (3), 340 – 357.
Nathan, P. J., Sitaram, G., Stough, C., Silberstein, R. B., & Sali, A. (2000). Serotonin, noradrenaline and cognitive
function: a preliminary investigation of the acute pharmacodynamic effects of a serotonin versus a serotoninand noradrenaline reuptake inhibitor. Behavioural Pharmacology, 11, 639 – 642.
Nathan, P. J., Stough, C., & Siteram, G. (2000). Serotonin and information processing: a pharmacodynamic study
on the effects of citalopram on cognitive and psychomotor function. Human Psychopharmacology: Clinicaland Experimental, 15, 306 – 307.
Nettelbeck, T. (1987). Inspection time and intelligence. In: P. A. Vernon (Ed.), Speed of information processing
and intelligence ( pp. 295 – 346). Norwood, NJ: Ablex.
Nettelbeck, T., & Young, R. (1989). Inspection time and intelligence in 6-year-old children. Personality and
Individual Differences, 10, 605 – 614.
Nettelbeck, T., & Young, R. (1990). Inspection time and intelligence in 7-year-old children. Personality and
Individual Differences, 11, 1283 – 1289.
Ollat, H. (1992). Dopaminergic insufficiency reflecting cerebral ageing: value of a dopaminergic agonist, piribedil.
Journal of Neurology, 239 (Suppl. 1), S13 – S16.
Parrott, A., & Deary, I. (1992). Scopolamine, cognition and dementia. Journal of Psychopharmacology, 6,
C. Stough et al. / Intelligence 29 (2001) 511–522
Phillips, J. M., & Brown, V. J. (1999). Reaction time performance following unilateral striatal dopamine depletion
and lesions of the subthalamic nucleus in the rat. European Journal of Neuroscience, 11 (3), 1003 – 1010.
Pomerleau, O. F., & Pomerleau, C. S. (1992). Nicotine replacement: a critical evaluation. New York: Haworth Press. Previc, F. H. (1999). Dopamine and the origins of human intelligence. Brain and Cognition, 41, 299 – 350. Pyman, Nathan, Thompson, & Stough (submitted for publication). The effects of 2 and 4 mg nicotine gum on the
speed of early information processing.
Robbins, T. W. (1998). Homology in behavioural pharmacology: an approach to animal models of human
cognition. Behavioural Pharmacology, 9 (7), 509 – 519.
Sarter, M., & Bruno, J. P. (1999). Abnormal regulation of corticopetal cholinergic neurons and impaired infor-
mation processing in neuropsychiatric disorders. Trends in Neurosciences, 22 (2), 67 – 74.
Sawaguchi, T., & Goldman-Rakic, P. (1997). D1 dopamine receptors in prefrontal cortex: involvement in working
Sawaguchi, T., Matsumura, M., & Kubota, K. (1990). Effects of dopamine antagonists on neural activity related to
a delayed response task in monkey’s prefrontal cortex. Journal of Neurophysiology, 63 (6), 1401 – 1412.
Smith, A. P., Wilson, S. J., Glue, P., & Nutt, D. J. (1992). The effects and after effects of the alpha2-adrenergic
antagonist idazoxan on mood, memory and attention in normal volunteers. Psychopharmacology, 6,376 – 381.
Stahl, S. M. (2000). Essential psychopharmacology (2nd ed.). London: Cambridge Univ. Press. Stitzel, J. A., Lu, Y., Jimenez, M., Tritto, T., & Collins, A. C. (2000). Genetic and pharmacological strategies
identify a behavioral function of neuronal nicotinic receptors. Behaviour Brain Research, 113 (1 – 2), 57 – 64.
Stough, C., Bates, T., Mangan, G. L., & Pellett, O. (1995). Smoking, string length and intelligence. Personality
and Individual Differences, 18, 75 – 79.
Stough, C., Brebner, J., Nettelbeck, T., Cooper, C. J., Bates, T. C., & Mangan, G. L. (1996). The relationship
between intelligence, personality and inspection time. British Journal of Psychology, 87, 255 – 268.
Stough, C., Mangan, G., Bates, T., Frank, N., Kerkin, B., & Pellett, O. (1995). Effects of nicotine on perceptual
speed? Psychopharmacology, 107, 305 – 310.
Stough, C., Mangan, G. L., Bates, T., & Pellett, O. (1994). Smoking and Raven IQ. Psychopharmacology, 106,
Stough, C., Sitaram, G., & Nathan, P. (submitted). Examining the serotonin and noradrenalin neurotransmitter
Stough, C., Singh, M., Lloyd, J., & Nathan, P. J. (submitted). Examining the effect of dopamine agonist on
Stough, C., Wilby, G., & Thompson, J. C. (submitted for publication). Transdermal nicotine administration and
Tani, Y., Saito, K., Imoto, M., & Ohno, T. (1998). Pharmacological characterization of nicotinic receptor-mediated
acetylcholine release in rat brain — an in vivo microdialysis study. European Journal of Pharmacology, 351(2), 181 – 188.
Thompson, J. C., Stough, C., Nathan, P. J., Ames, D., & Ritchie, C. (2000). Effects of the nicotinic antagonist
mecamylamine on inspection time. Psychopharmacology, 150 (1), 117 – 119.
Vickers, D., Nettelbeck, T., & Willson, R. J. (1972). Perceptual indices of performance: the measurement of
‘‘inspection time’’ and ‘‘noise’’ in the visual system. Perception, 1, 263 – 295.
Volkow, N., Gur, R., Wang, G., & Fowler, J. (1998). Association between decline in brain dopamine activity with
age and cognitive and motor impairment in health individuals. American Journal of Psychiatry, 155 (3),344 – 348.
Warburton, D. M. (1992). Nicotine as a cognitive enhancer. Progress in Neuro-Psychopharmacology & Biological
Waterham, I., Thompson, J. C., Nathan, P. J., & Stough, C. (submitted for publication). Whitehouse, P. J., & Kalaria, R. N. (1995). Nicotinic receptors and neurodegenerative dementing diseases: basic
research and clinical implications. Alzheimer Disease and Associated Disorders, 2 (Suppl. 9), 3 – 5.
aNortheastern Ohio Universities College of Medicine, 4209 State Route 44, PO Box 95,bDivision of Sports Medicine, Akron Children’s Hospital, Sports Medicine Center,388 South Main Street, Suite 207, Akron, OH 44311, USAThe ‘‘win at all costs’’ mentality fuels athletes to seek performance-enhancing substances, such as anabolic-androgenic steroids (AASs), togain an advantage over th
TO: Columbus Area EMS District Board Members FROM: Werner Biedermann DATE: July 10, 2013 SUBJECT: June 2013 Monthly Report Lifestar responded to 54 emergency 911 calls in June. We also performed a paramedic intercept for Randolph EMS and one fire standby for Fall River Fire Department. Below is the call breakdown by municipality. Columbus Contract Run Statistics for 2013 JA

 Examining neurochemical determinants of inspection time
Con Stougha,*, J.C. Thompsonb, T.C. Batesc, P.J. Nathand
aNeuropsychology Laboratory, BSEE, Swinburne University, P.O. Box 218, Hawthorn, VIC 3122, Australia
bBrain Sciences Institute, Swinburne University, Hawthorn, VIC 3122, Australia
cDepartment of Psychology, Macquarie University, North Ryde, NSW, Australia
dNeuropharmacology Laboratory, Swinburne University, Hawthorn, VIC 3122, Australia
Received 13 March 2001; accepted 13 March 2001
Inspection time (IT), an information-processing correlate of psychometric intelligence, has been
extensively studied. Previous research has shown that IT is a reliable correlate of psychometricintelligence across different developmental periods, mirroring developmental trends of fluidintelligence. Despite this extensive previous literature, very little is known about the biological basisof IT. In the present review, we discuss recent results from our laboratories examining theneurochemical determinants of IT. In this review, we outline the significance of several studies inwhich performance on the IT task is measured before and after modulating key human central nervoussystem (CNS) neurotransmitters and receptor systems (e.g., cholinergic, serotonergic, noradrenergic,and dopaminergic systems). The results of these studies indicate a primarily cholinergic basis for IT,although other aspects of psychometric intelligence may have serotonergic and dopaminergicdeterminants in addition to a cholinergic basis. The results are consistent with data reportingcholinergic depletion and impaired IT performance in dementia of the Alzheimer’s type (DAT).
Examining neurochemical determinants of inspection time
Con Stougha,*, J.C. Thompsonb, T.C. Batesc, P.J. Nathand
aNeuropsychology Laboratory, BSEE, Swinburne University, P.O. Box 218, Hawthorn, VIC 3122, Australia
bBrain Sciences Institute, Swinburne University, Hawthorn, VIC 3122, Australia
cDepartment of Psychology, Macquarie University, North Ryde, NSW, Australia
dNeuropharmacology Laboratory, Swinburne University, Hawthorn, VIC 3122, Australia
Received 13 March 2001; accepted 13 March 2001
Inspection time (IT), an information-processing correlate of psychometric intelligence, has been
extensively studied. Previous research has shown that IT is a reliable correlate of psychometricintelligence across different developmental periods, mirroring developmental trends of fluidintelligence. Despite this extensive previous literature, very little is known about the biological basisof IT. In the present review, we discuss recent results from our laboratories examining theneurochemical determinants of IT. In this review, we outline the significance of several studies inwhich performance on the IT task is measured before and after modulating key human central nervoussystem (CNS) neurotransmitters and receptor systems (e.g., cholinergic, serotonergic, noradrenergic,and dopaminergic systems). The results of these studies indicate a primarily cholinergic basis for IT,although other aspects of psychometric intelligence may have serotonergic and dopaminergicdeterminants in addition to a cholinergic basis. The results are consistent with data reportingcholinergic depletion and impaired IT performance in dementia of the Alzheimer’s type (DAT).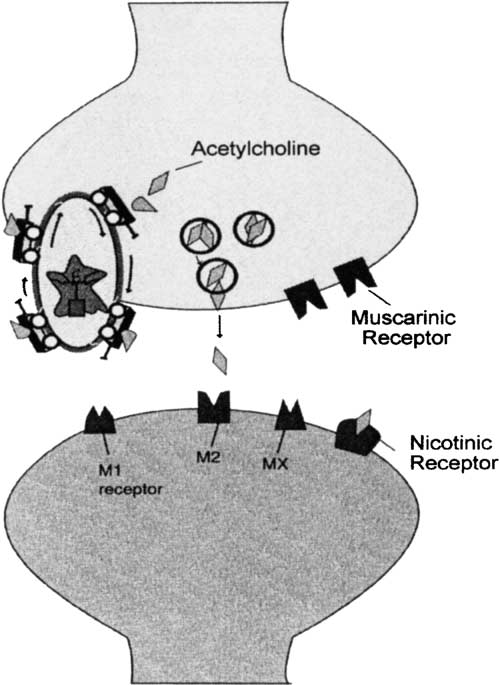 C. Stough et al. / Intelligence 29 (2001) 511–522
healthy participants. We outline our theory and empirical support for a predominantlycholinergic basis for IT.
C. Stough et al. / Intelligence 29 (2001) 511–522
healthy participants. We outline our theory and empirical support for a predominantlycholinergic basis for IT.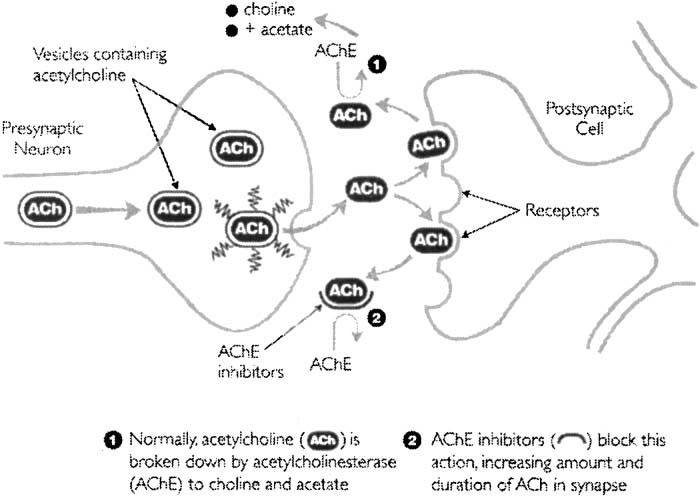 C. Stough et al. / Intelligence 29 (2001) 511–522
(i.e., presynaptic release). This increase in Ach increases the probability of the neuro-transmitter, finding a suitable postsynaptic receptor (in the case of the cholinergic system, thisis predominantly muscarinic receptors but there is also limited postsynaptic nicotinicreceptors). Fig. 1 also illustrates the key components of the cholinergic system includingneurotransmitter (Ach), pre- and postsynaptic receptors (nicotinic and muscarinic), andenzyme that metabolises Ach. In Fig. 1, Ach is released from presynaptic vesicles (middleof Fig. 1). It should probably also be pointed out here that the neurochemical approach thatwe have employed is to systematically manipulate the various elements of this system and toobserve the effect on IT performance. In all studies, we have employed placebo-controlleddouble-blind methodologies. Fig. 2 illustratively describes the mechanism by which theneurotransmitter Ach and acetylcholinesterase inhibitor act within this system. This is a keyelement of this system and will be discussed in more detail later.
C. Stough et al. / Intelligence 29 (2001) 511–522
(i.e., presynaptic release). This increase in Ach increases the probability of the neuro-transmitter, finding a suitable postsynaptic receptor (in the case of the cholinergic system, thisis predominantly muscarinic receptors but there is also limited postsynaptic nicotinicreceptors). Fig. 1 also illustrates the key components of the cholinergic system includingneurotransmitter (Ach), pre- and postsynaptic receptors (nicotinic and muscarinic), andenzyme that metabolises Ach. In Fig. 1, Ach is released from presynaptic vesicles (middleof Fig. 1). It should probably also be pointed out here that the neurochemical approach thatwe have employed is to systematically manipulate the various elements of this system and toobserve the effect on IT performance. In all studies, we have employed placebo-controlleddouble-blind methodologies. Fig. 2 illustratively describes the mechanism by which theneurotransmitter Ach and acetylcholinesterase inhibitor act within this system. This is a keyelement of this system and will be discussed in more detail later.